

Short Direct Repeats in the 3' Untranslated Region Are Involved in Subgenomic Flaviviral RNA Production
- PMID: 31896596
- PMCID: PMC7158718
- DOI: 10.1128/JVI.01175-19
Short Direct Repeats in the 3' Untranslated Region Are Involved in Subgenomic Flaviviral RNA Production
Abstract
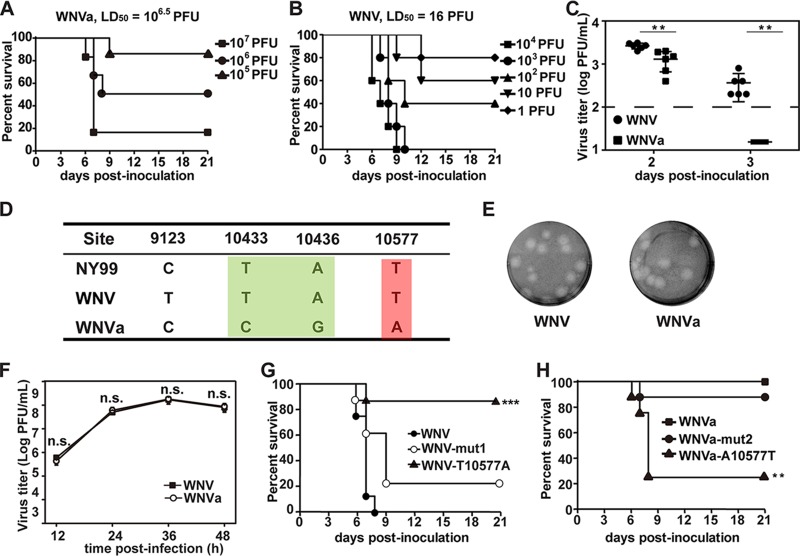
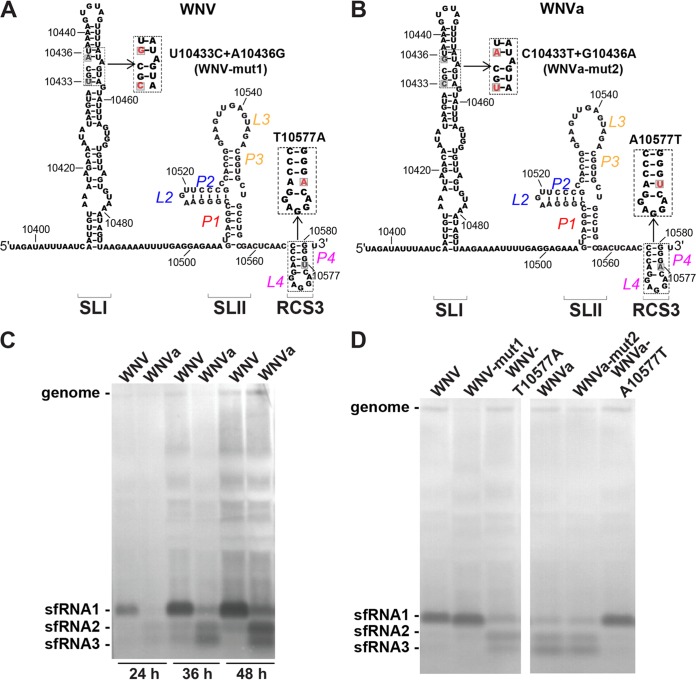
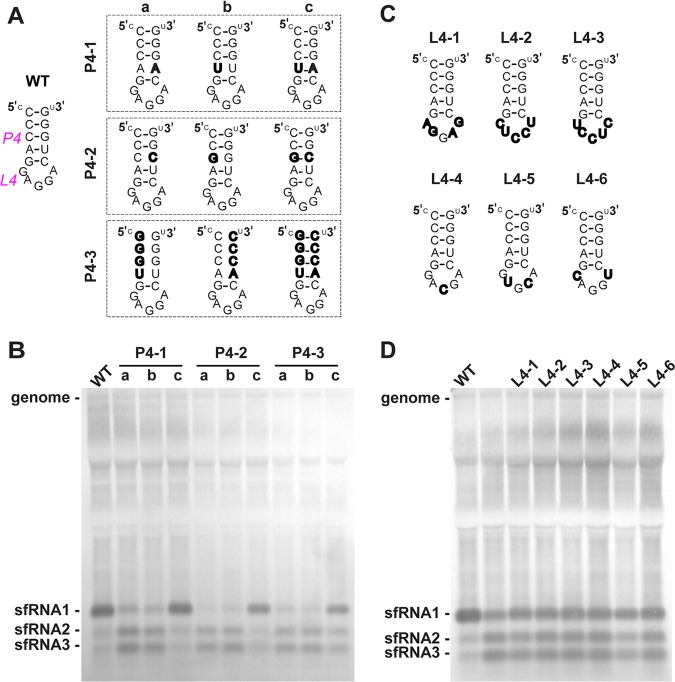
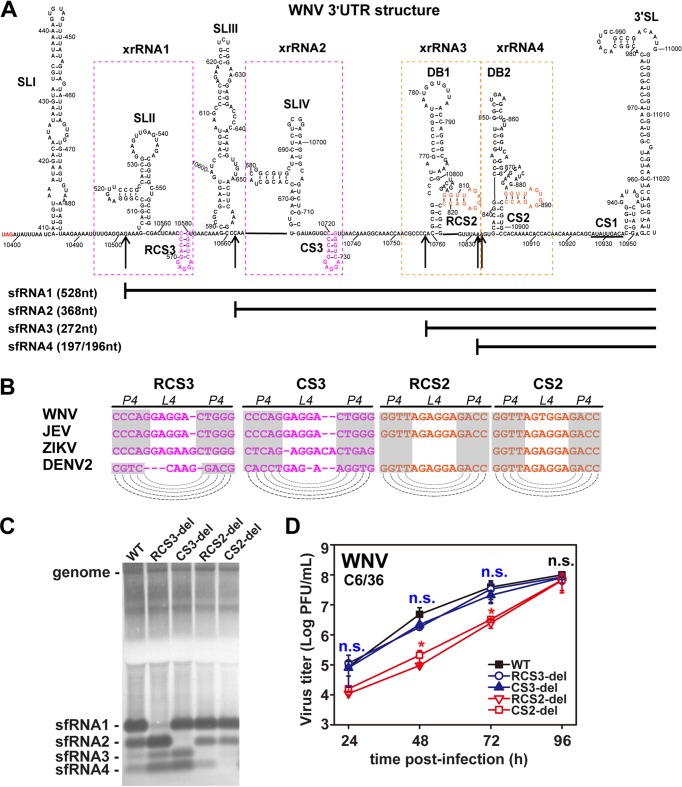
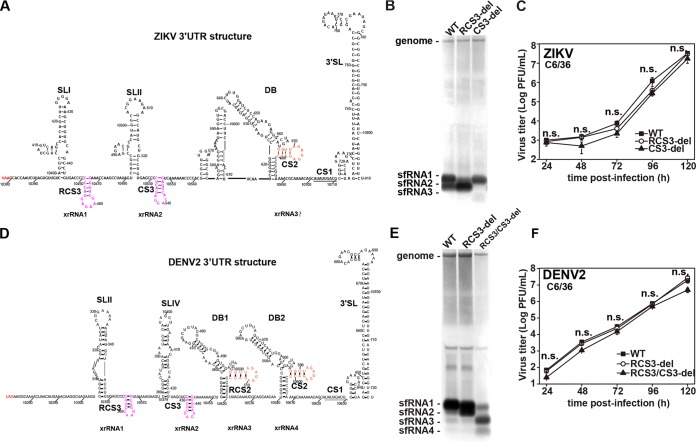
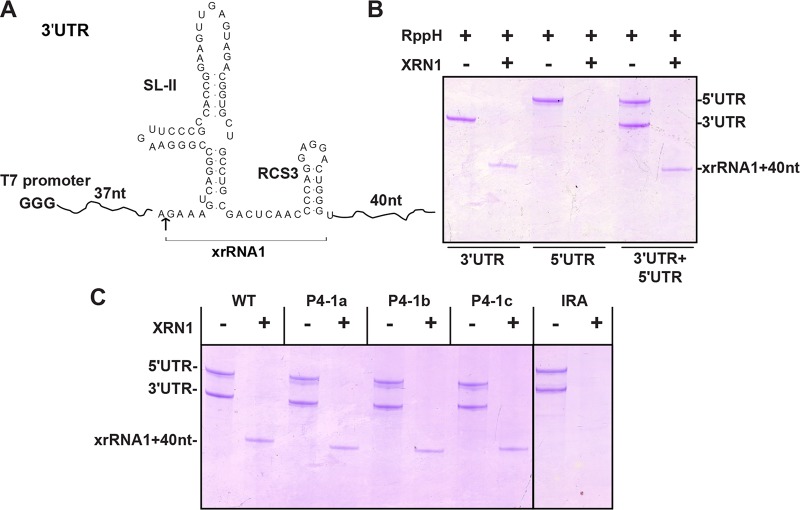
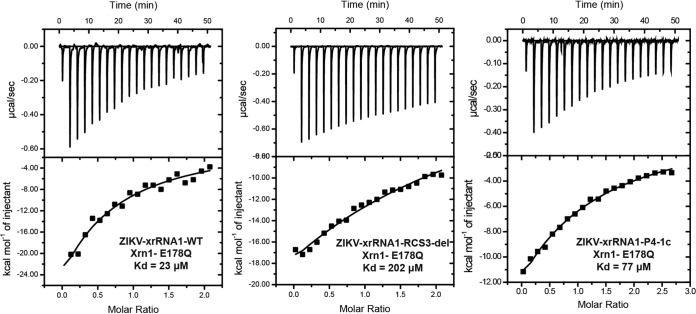
Mosquito-borne flaviviruses consist of a positive-sense genome RNA flanked by the untranslated regions (UTRs). There is a panel of highly complex RNA structures in the UTRs with critical functions. For instance, Xrn1-resistant RNAs (xrRNAs) halt Xrn1 digestion, leading to the production of subgenomic flaviviral RNA (sfRNA). Conserved short direct repeats (DRs), also known as conserved sequences (CS) and repeated conserved sequences (RCS), have been identified as being among the RNA elements locating downstream of xrRNAs, but their biological function remains unknown. In this study, we revealed that the specific DRs are involved in the production of specific sfRNAs in both mammalian and mosquito cells. Biochemical assays and structural remodeling demonstrate that the base pairings in the stem of these DRs control sfRNA formation by maintaining the binding affinity of the corresponding xrRNAs to Xrn1. On the basis of these findings, we propose that DRs functions like a bracket holding the Xrn1-xrRNA complex for sfRNA formation.IMPORTANCE Flaviviruses include many important human pathogens. The production of subgenomic flaviviral RNAs (sfRNAs) is important for viral pathogenicity as a common feature of flaviviruses. sfRNAs are formed through the incomplete degradation of viral genomic RNA by the cytoplasmic 5'-3' exoribonuclease Xrn1 halted at the Xrn1-resistant RNA (xrRNA) structures within the 3'-UTR. The 3'-UTRs of the flavivirus genome also contain distinct short direct repeats (DRs), such as RCS3, CS3, RCS2, and CS2. However, the biological functions of these ancient primary DR sequences remain largely unknown. Here, we found that DR sequences are involved in sfRNA formation and viral virulence and provide novel targets for the rational design of live attenuated flavivirus vaccine.
Keywords: Flavivirus; West Nile virus; direct repeats; sfRNA.
Copyright © 2020 American Society for Microbiology.
Figures
References
-
- Filomatori CV, Carballeda JM, Villordo SM, Aguirre S, Pallares HM, Maestre AM, Sanchez-Vargas I, Blair CD, Fabri C, Morales MA, Fernandez-Sesma A, Gamarnik AV. 2017. Dengue virus genomic variation associated with mosquito adaptation defines the pattern of viral non-coding RNAs and fitness in human cells. PLoS Pathog 13:e1006265. doi:10.1371/journal.ppat.1006265. - DOI - PMC - PubMed
-
- Manokaran G, Finol E, Wang C, Gunaratne J, Bahl J, Ong EZ, Tan HC, Sessions OM, Ward AM, Gubler DJ, Harris E, Garcia-Blanco MA, Ooi EE. 2015. Dengue subgenomic RNA binds TRIM25 to inhibit interferon expression for epidemiological fitness. Science 350:217–221. doi:10.1126/science.aab3369. - DOI - PMC - PubMed
-
- Pijlman GP, Funk A, Kondratieva N, Leung J, Torres S, van der Aa L, Liu WJ, Palmenberg AC, Shi PY, Hall RA, Khromykh AA. 2008. A highly structured, nuclease-resistant, noncoding RNA produced by flaviviruses is required for pathogenicity. Cell Host Microbe 4:579–591. doi:10.1016/j.chom.2008.10.007. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
