

Retinoic acid degradation shapes zonal development of vestibular organs and sensitivity to transient linear accelerations
- PMID: 31896743
- PMCID: PMC6940366
- DOI: 10.1038/s41467-019-13710-4
Retinoic acid degradation shapes zonal development of vestibular organs and sensitivity to transient linear accelerations
Abstract
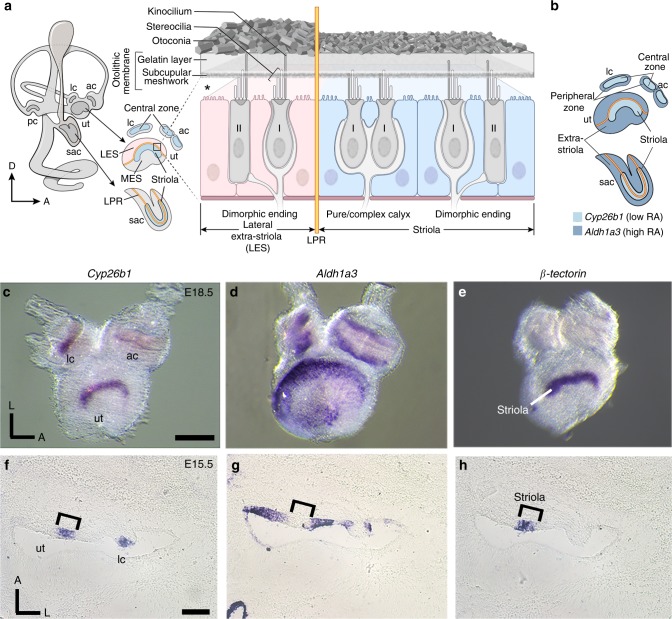
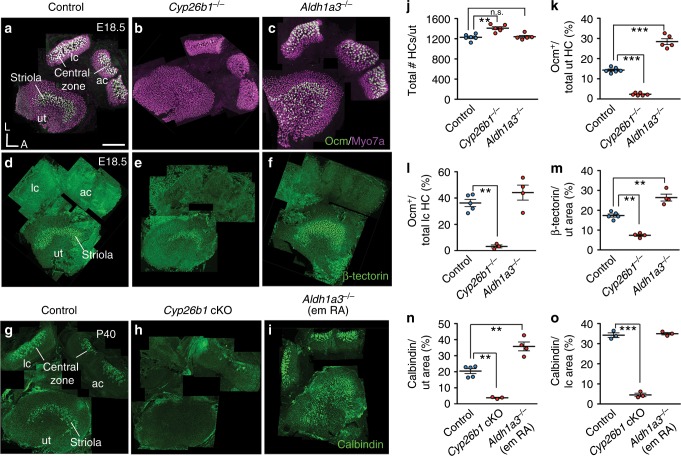
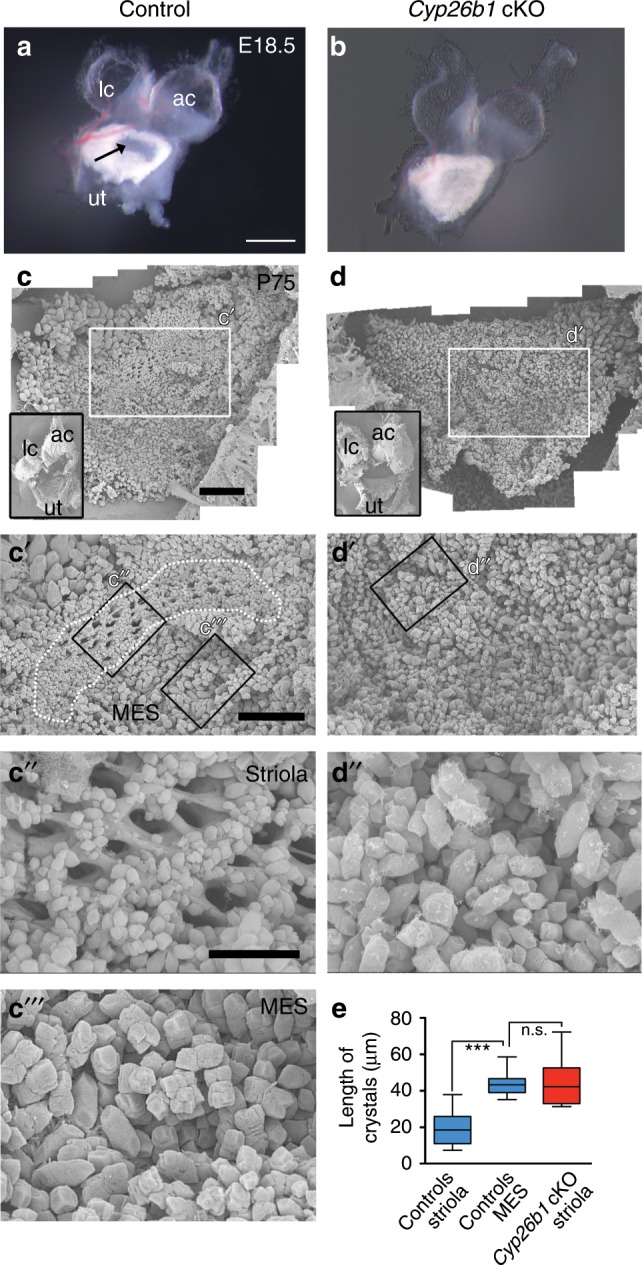
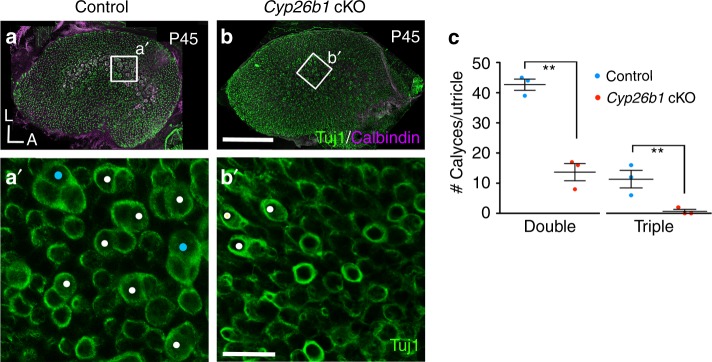
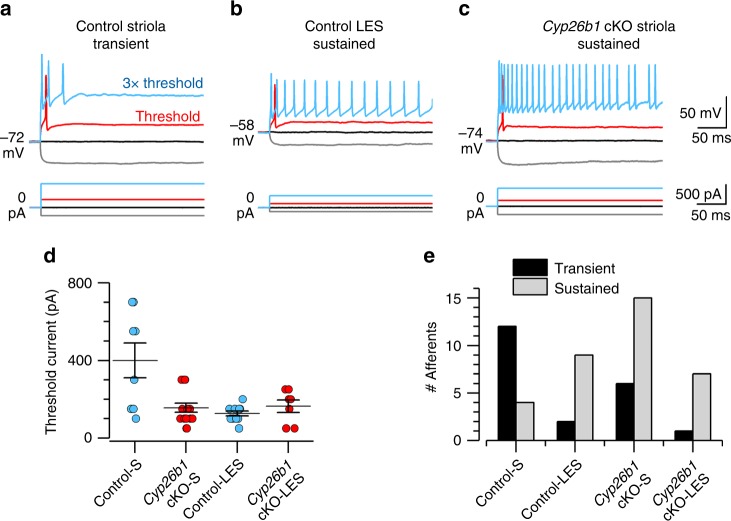
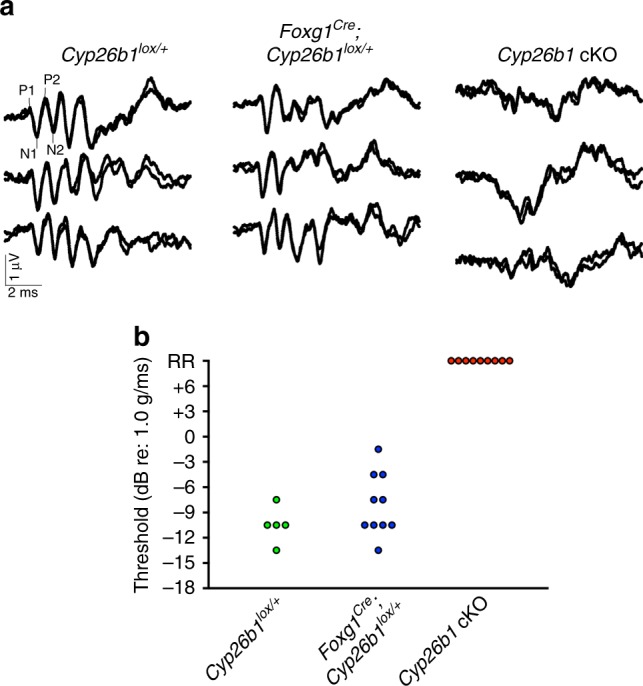
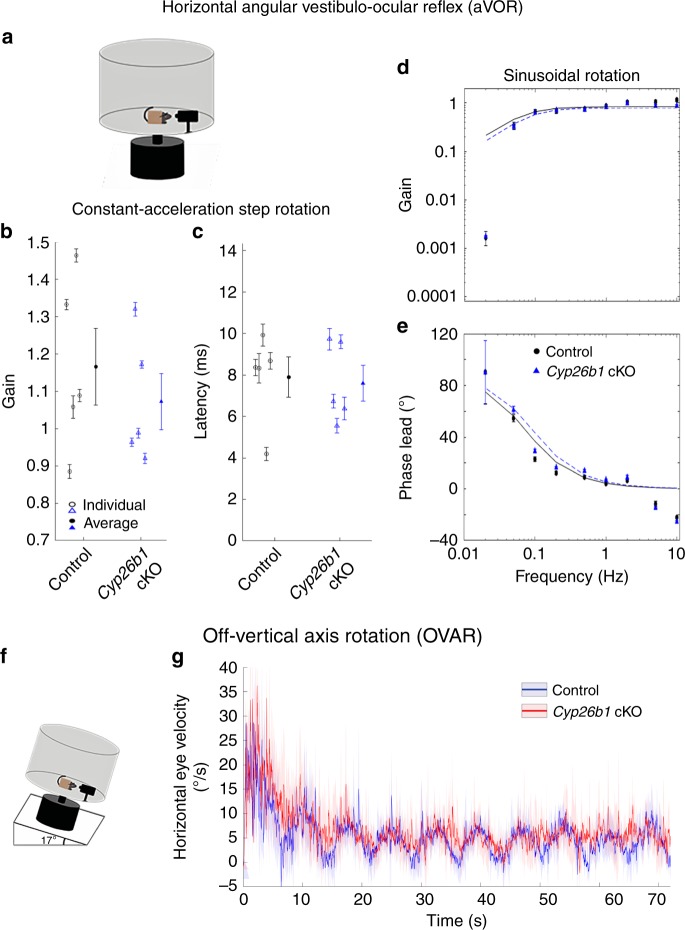
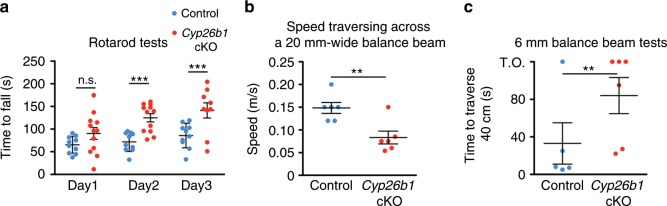
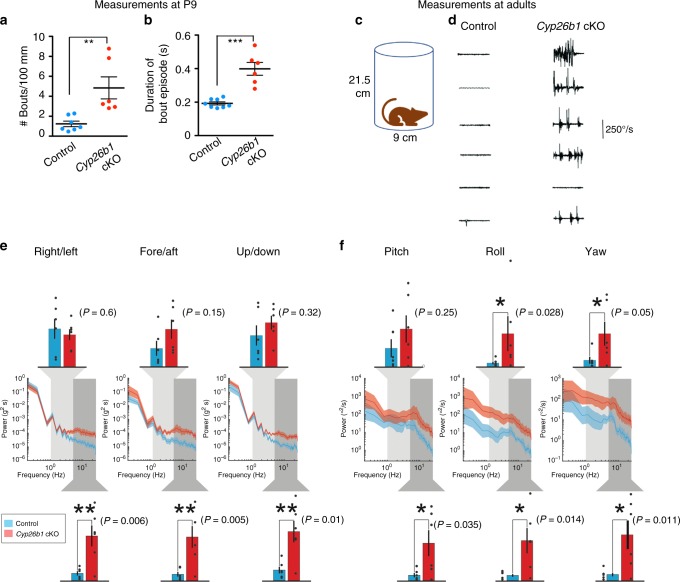
Each vestibular sensory epithelium in the inner ear is divided morphologically and physiologically into two zones, called the striola and extrastriola in otolith organ maculae, and the central and peripheral zones in semicircular canal cristae. We found that formation of striolar/central zones during embryogenesis requires Cytochrome P450 26b1 (Cyp26b1)-mediated degradation of retinoic acid (RA). In Cyp26b1 conditional knockout mice, formation of striolar/central zones is compromised, such that they resemble extrastriolar/peripheral zones in multiple features. Mutants have deficient vestibular evoked potential (VsEP) responses to jerk stimuli, head tremor and deficits in balance beam tests that are consistent with abnormal vestibular input, but normal vestibulo-ocular reflexes and apparently normal motor performance during swimming. Thus, degradation of RA during embryogenesis is required for formation of highly specialized regions of the vestibular sensory epithelia with specific functions in detecting head motions.
Conflict of interest statement
The authors have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
