

Continuous endoglin (CD105) overexpression disrupts angiogenesis and facilitates tumor cell metastasis
- PMID: 31897911
- PMCID: PMC7160077
- DOI: 10.1007/s10456-019-09703-y
Continuous endoglin (CD105) overexpression disrupts angiogenesis and facilitates tumor cell metastasis
Abstract
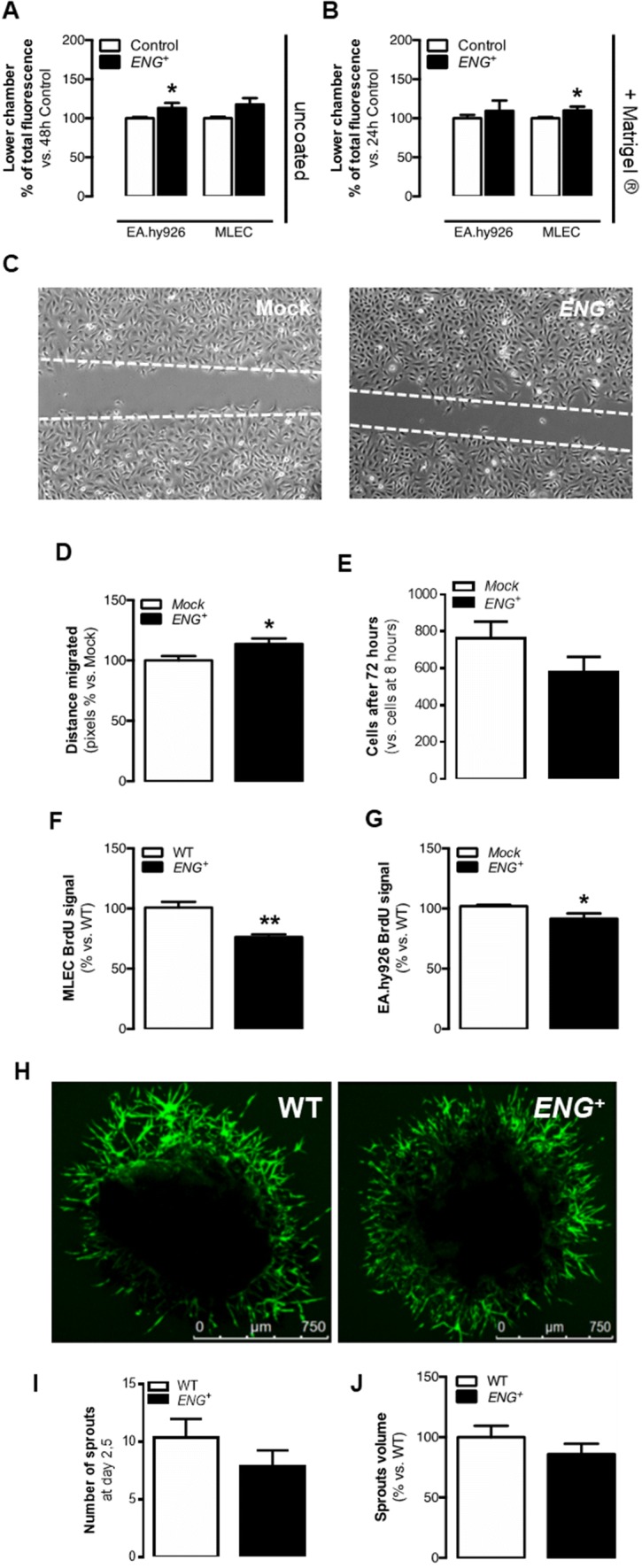
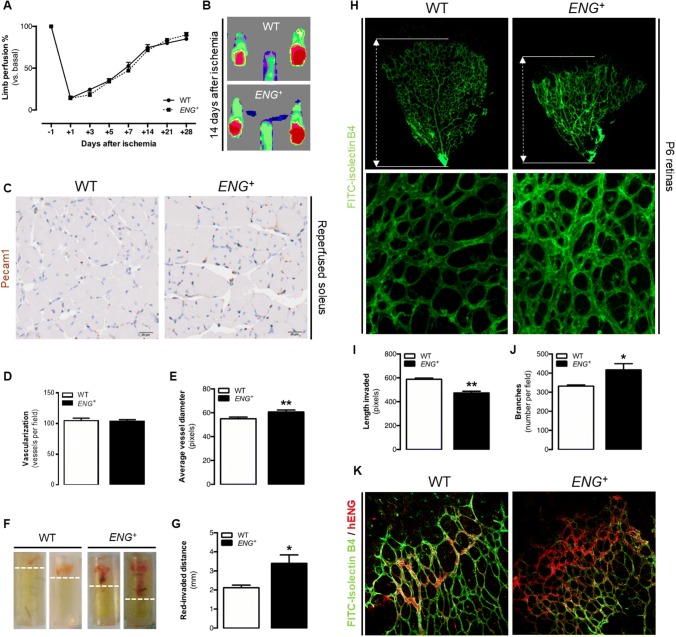
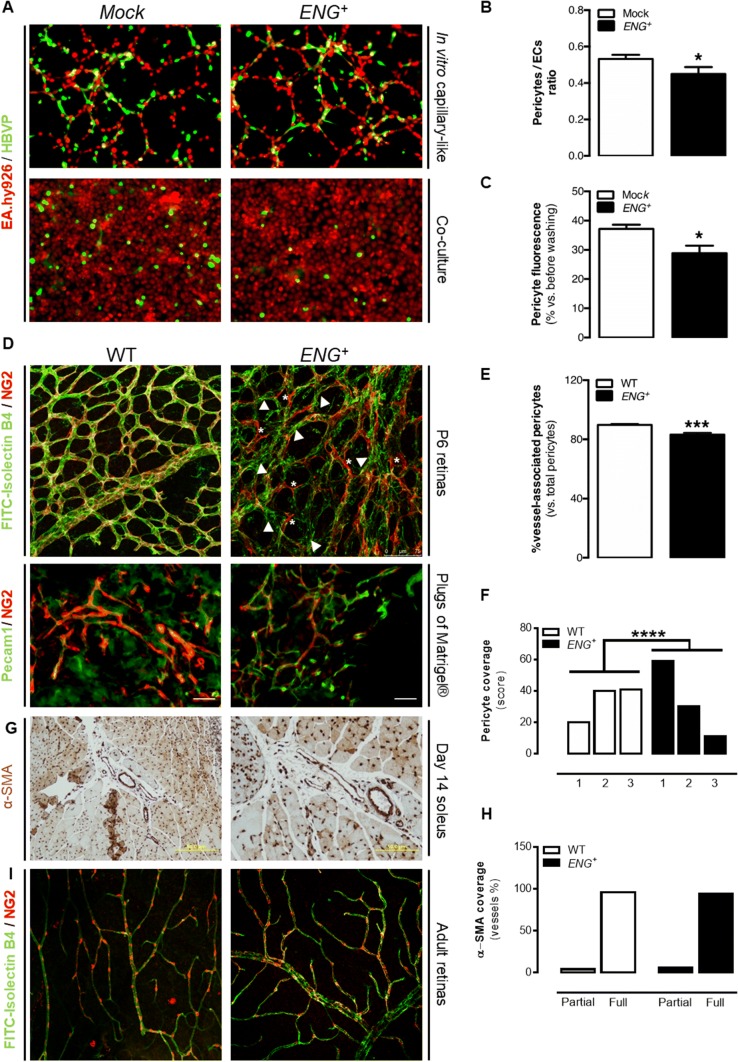
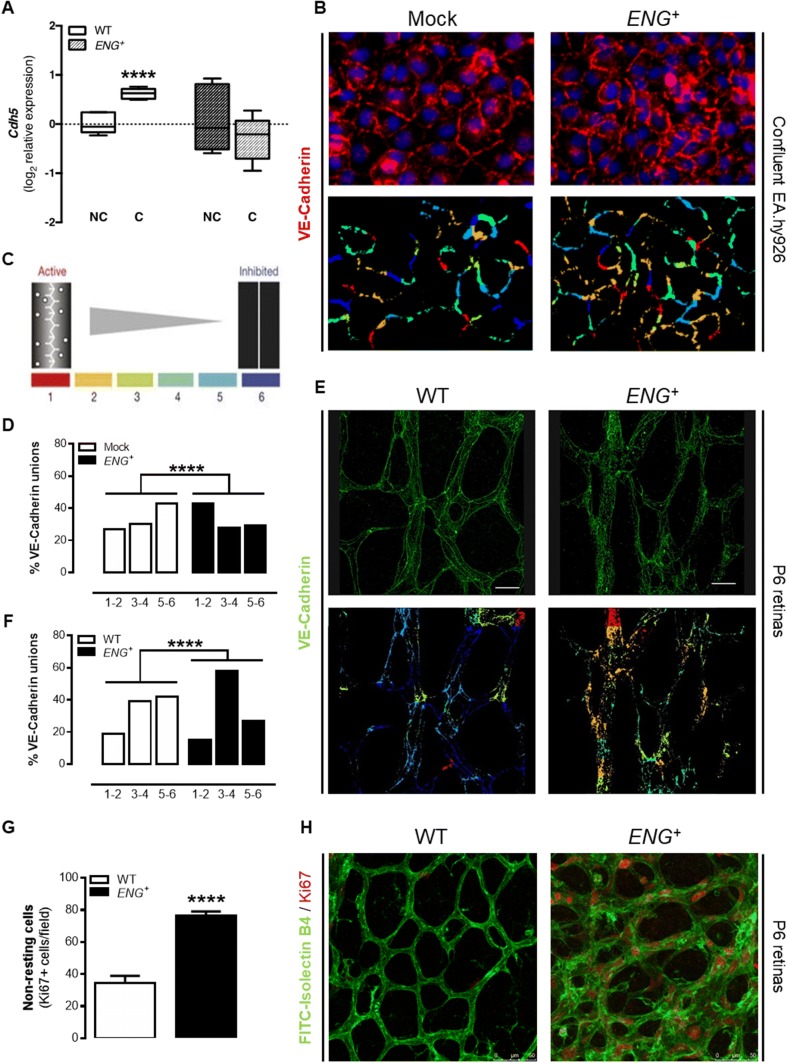
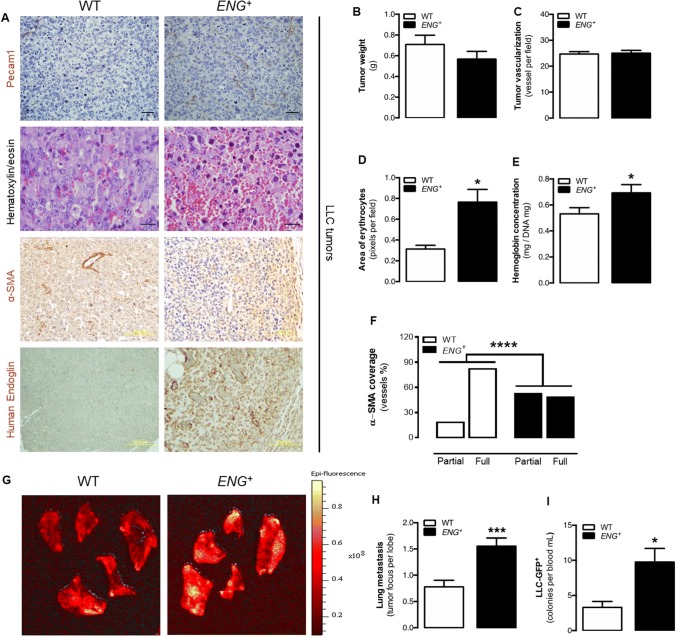
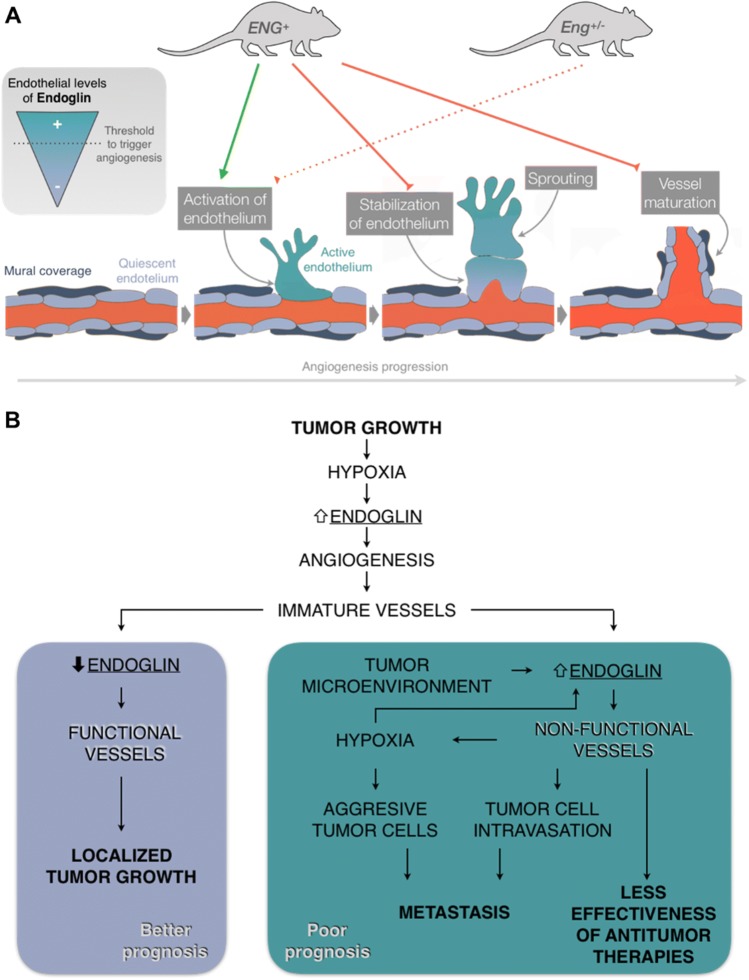
Endoglin (CD105) is an auxiliary receptor for members of the TFG-β superfamily. Whereas it has been demonstrated that the deficiency of endoglin leads to minor and defective angiogenesis, little is known about the effect of its increased expression, characteristic of several types of cancer. Angiogenesis is essential for tumor growth, so high levels of proangiogenic molecules, such as endoglin, are supposed to be related to greater tumor growth leading to a poor cancer prognosis. However, we demonstrate here that endoglin overexpression do not stimulate sprouting or vascularization in several in vitro and in vivo models. Instead, steady endoglin overexpression keep endothelial cells in an active phenotype that results in an impairment of the correct stabilization of the endothelium and the recruitment of mural cells. In a context of continuous enhanced angiogenesis, such as in tumors, endoglin overexpression gives rise to altered vessels with an incomplete mural coverage that permit the extravasation of blood. Moreover, these alterations allow the intravasation of tumor cells, the subsequent development of metastases and, thus, a worse cancer prognosis.
Keywords: Angiogenesis; CD105; Cancer; Endoglin; Metastasis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Expression of Endoglin and Vascular Endothelial Growth Factor as Prognostic Markers in Experimental Colorectal Cancer.Anticancer Res. 2016 Aug;36(8):3953-9. Anticancer Res. 2016. PMID: 27466499
-
CD105 is important for angiogenesis: evidence and potential applications.FASEB J. 2003 Jun;17(9):984-92. doi: 10.1096/fj.02-0634rev. FASEB J. 2003. PMID: 12773481 Review.
-
Compound genetically engineered mouse models of cancer reveal dual targeting of ALK1 and endoglin as a synergistic opportunity to impinge on angiogenic TGF-β signaling.Oncotarget. 2016 Dec 20;7(51):84314-84325. doi: 10.18632/oncotarget.12604. Oncotarget. 2016. PMID: 27741515 Free PMC article.
-
Endoglin interacts with VEGFR2 to promote angiogenesis.FASEB J. 2018 Jun;32(6):2934-2949. doi: 10.1096/fj.201700867RR. Epub 2018 Jan 11. FASEB J. 2018. PMID: 29401587
-
Endoglin (CD105): a review of its role in angiogenesis and tumor diagnosis, progression and therapy.Anticancer Res. 2011 Jun;31(6):2283-90. Anticancer Res. 2011. PMID: 21737653 Review.
Cited by
-
CD105 expression is associated with invasive capacity in ovarian cancer and promotes invasiveness by inhibiting NDRG1 and regulating the epithelial-mesenchymal transition.Am J Transl Res. 2021 Nov 15;13(11):12461-12479. eCollection 2021. Am J Transl Res. 2021. PMID: 34956466 Free PMC article.
-
NRP1 interacts with endoglin and VEGFR2 to modulate VEGF signaling and endothelial cell sprouting.Commun Biol. 2024 Jan 19;7(1):112. doi: 10.1038/s42003-024-05798-2. Commun Biol. 2024. PMID: 38242992 Free PMC article.
-
Targeting CD44 and other pleiotropic co-receptors as a means for broad inhibition of tumor growth and metastasis.Clin Exp Metastasis. 2024 Oct;41(5):599-611. doi: 10.1007/s10585-024-10292-4. Epub 2024 May 18. Clin Exp Metastasis. 2024. PMID: 38761292 Free PMC article. Review.
-
Hot and Cold Tumors: Is Endoglin (CD105) a Potential Target for Vessel Normalization?Cancers (Basel). 2021 Mar 28;13(7):1552. doi: 10.3390/cancers13071552. Cancers (Basel). 2021. PMID: 33800564 Free PMC article. Review.
-
Curcuminoids Inhibit Angiogenic Behaviors of Human Umbilical Vein Endothelial Cells via Endoglin/Smad1 Signaling.Int J Mol Sci. 2022 Mar 31;23(7):3889. doi: 10.3390/ijms23073889. Int J Mol Sci. 2022. PMID: 35409247 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- SAF2010-15881/Ministerio de Economía y Competitividad/International
- SAF2013-45784-R/Ministerio de Economía y Competitividad/International
- RD06/0016/0013/Instituto de Salud Carlos III/International
- RD12/0021/0032/Instituto de Salud Carlos III/International
- RD016/0009/0025/Instituto de Salud Carlos III/International
