

IL-17 signaling in steatotic hepatocytes and macrophages promotes hepatocellular carcinoma in alcohol-related liver disease
- PMID: 31899206
- PMCID: PMC7167339
- DOI: 10.1016/j.jhep.2019.12.016
IL-17 signaling in steatotic hepatocytes and macrophages promotes hepatocellular carcinoma in alcohol-related liver disease
Abstract
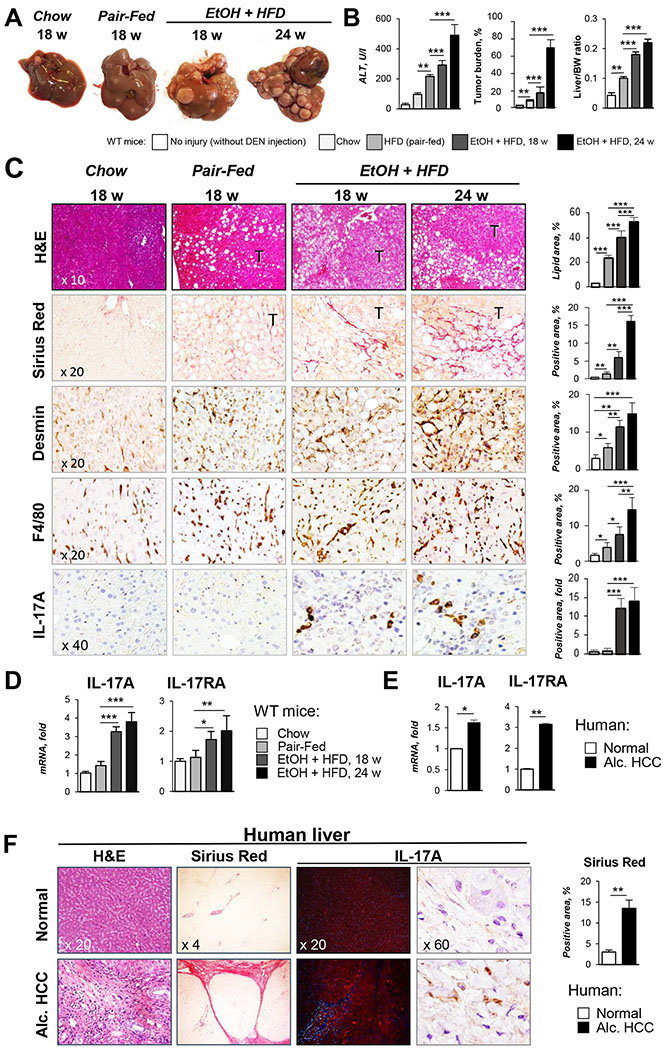
Background & aims: Chronic alcohol consumption is a leading risk factor for the development of hepatocellular carcinoma (HCC), which is associated with a marked increase in hepatic expression of pro-inflammatory IL-17A and its receptor IL-17RA.
Methods: Genetic deletion and pharmacological blocking were used to characterize the role of IL-17A/IL-17RA signaling in the pathogenesis of HCC in mouse models and human specimens.
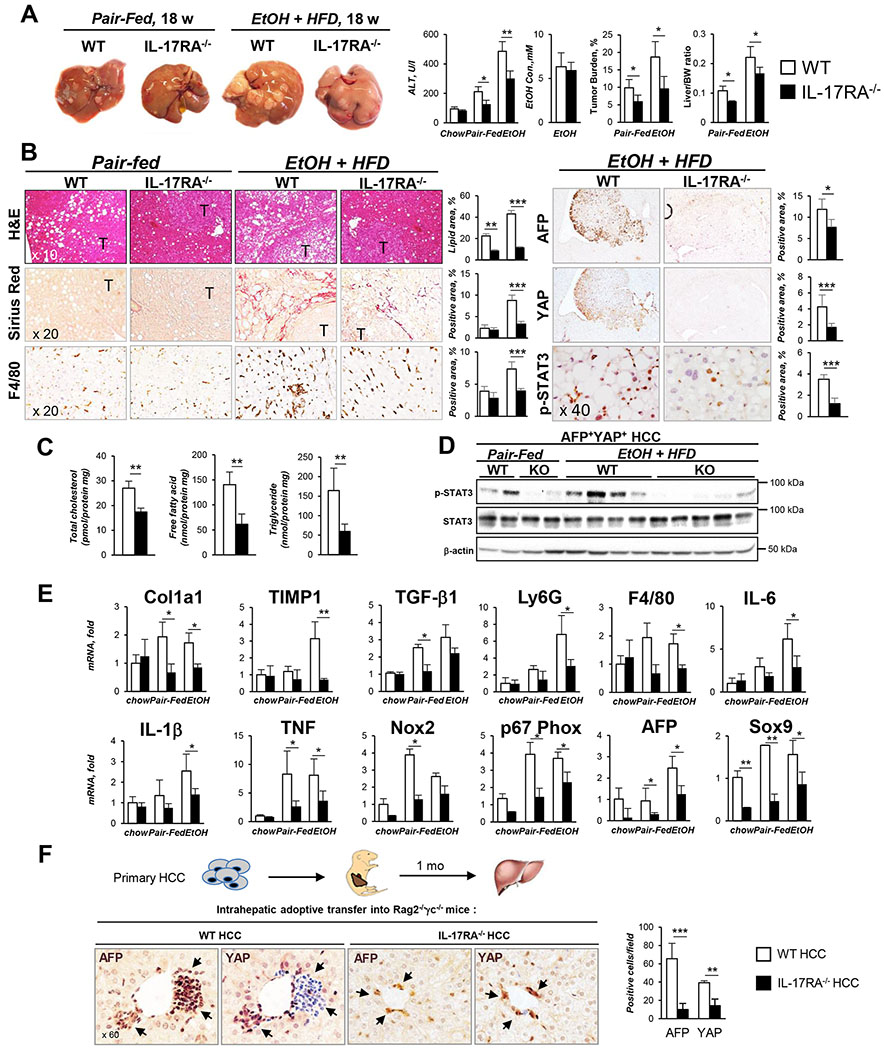
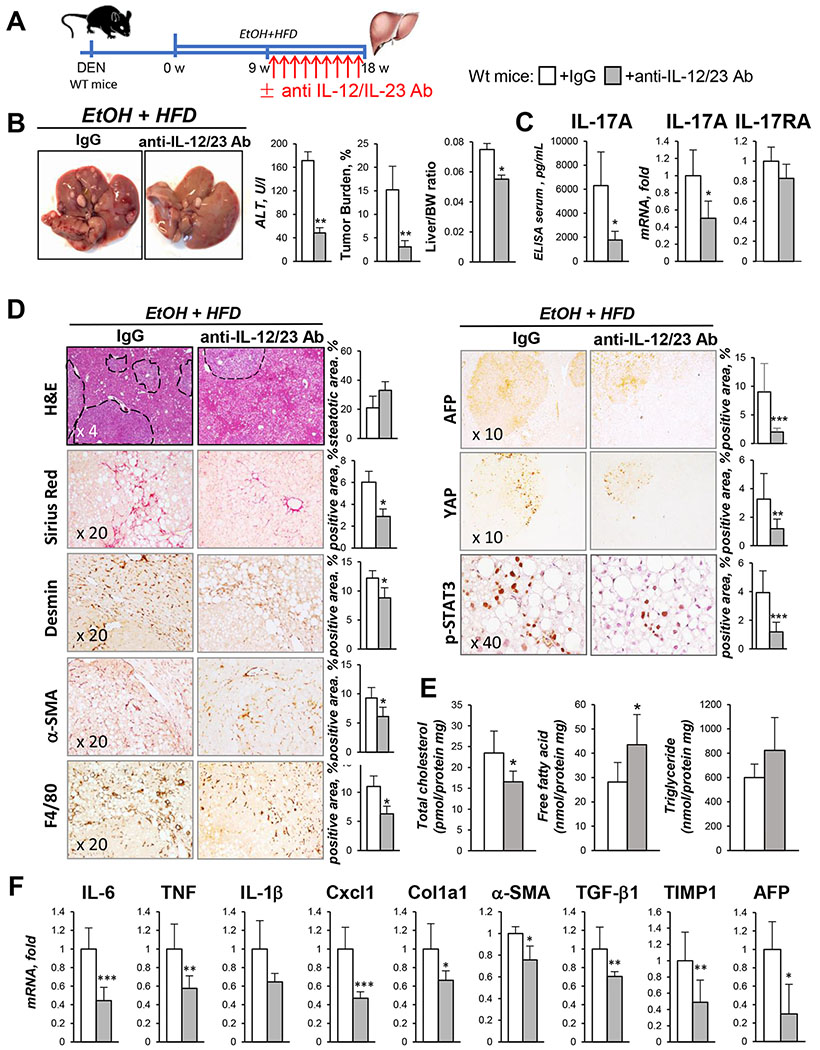
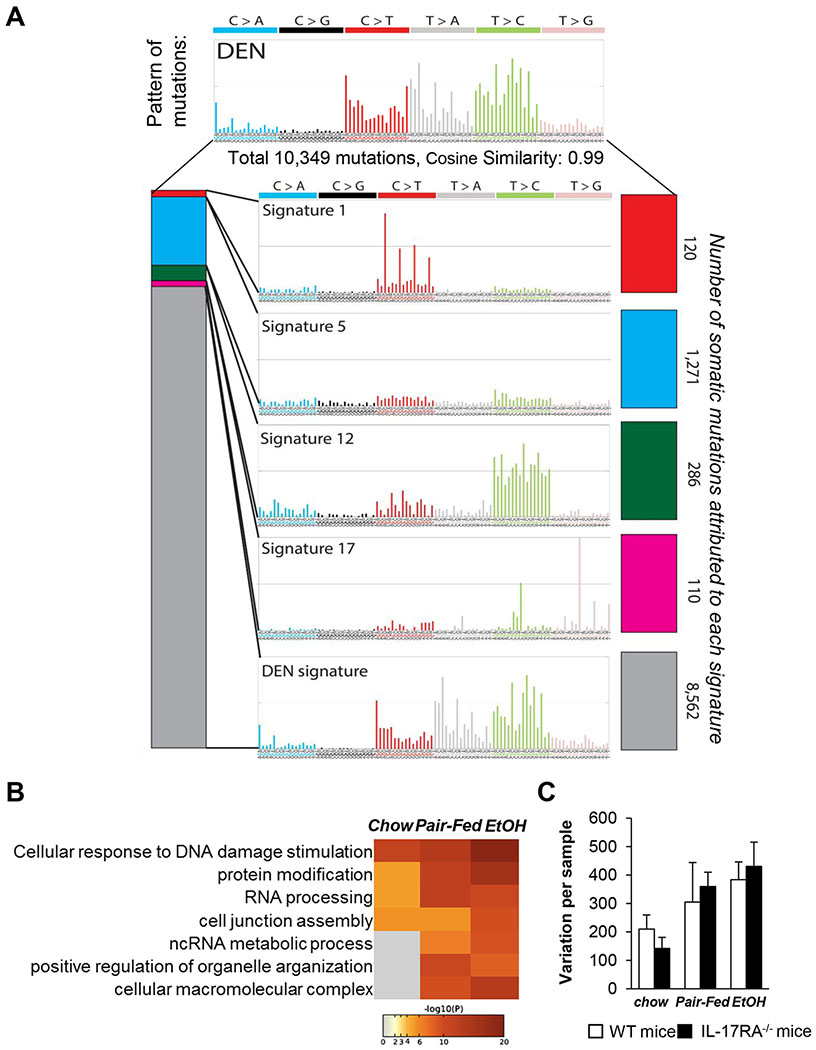
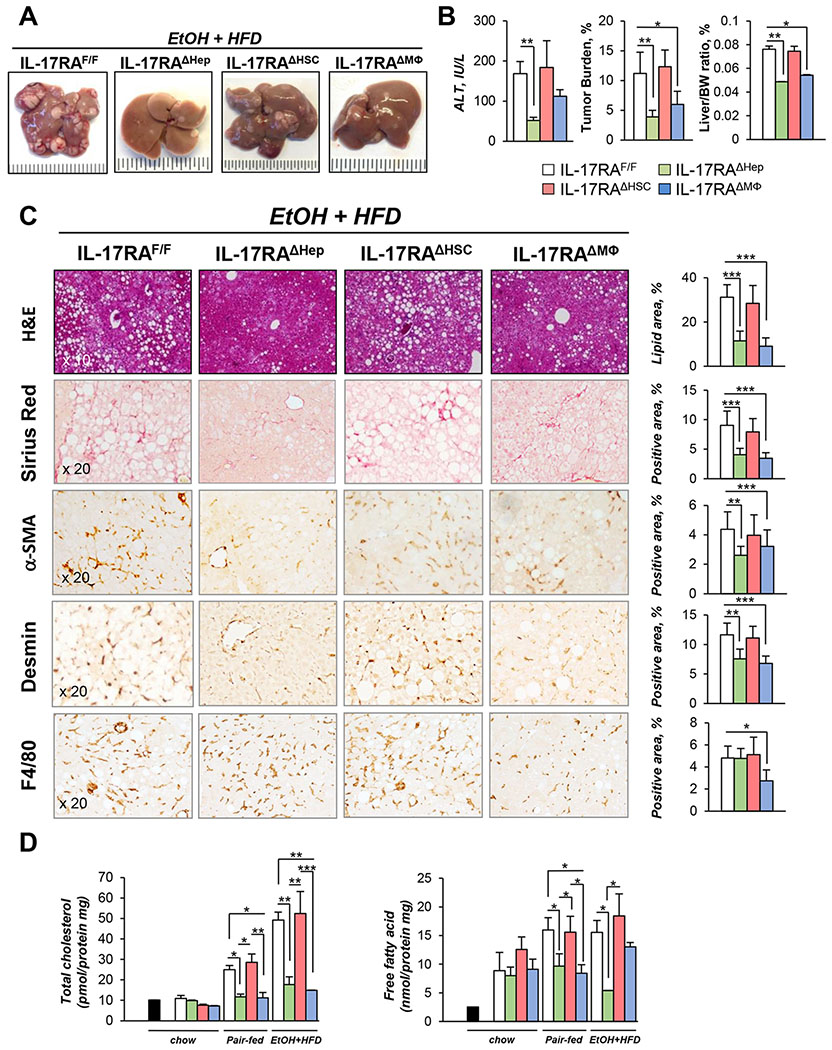
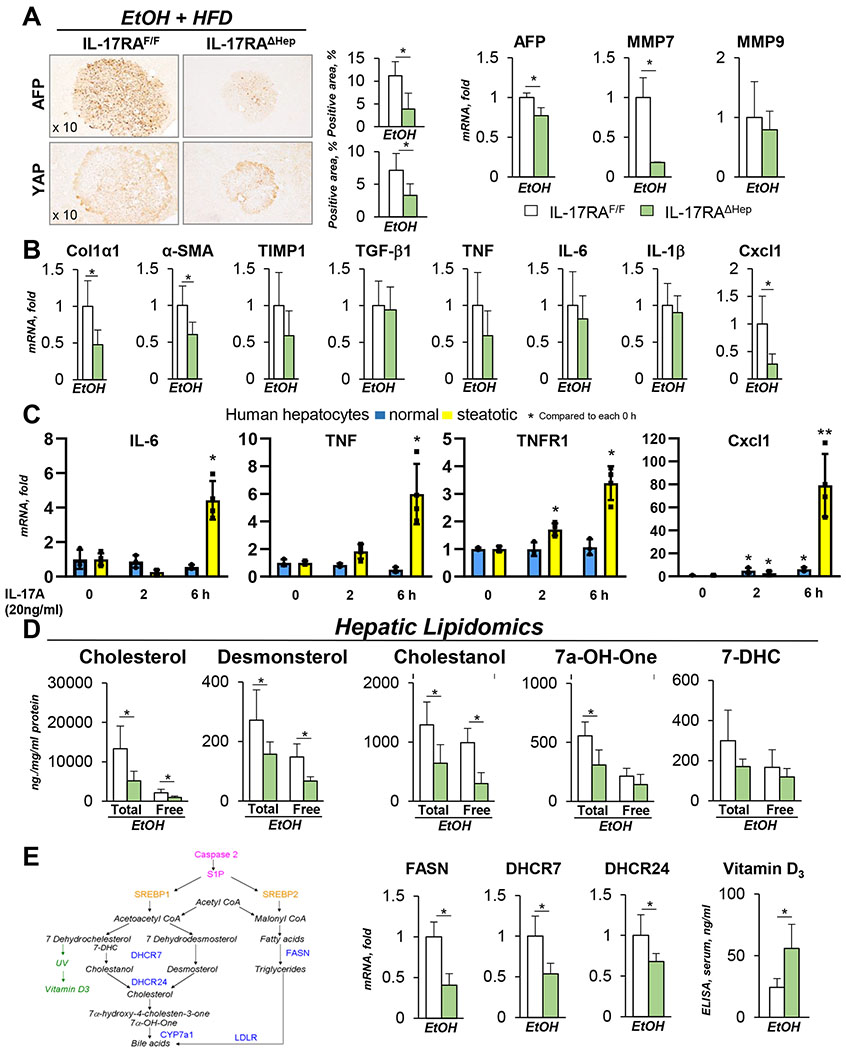
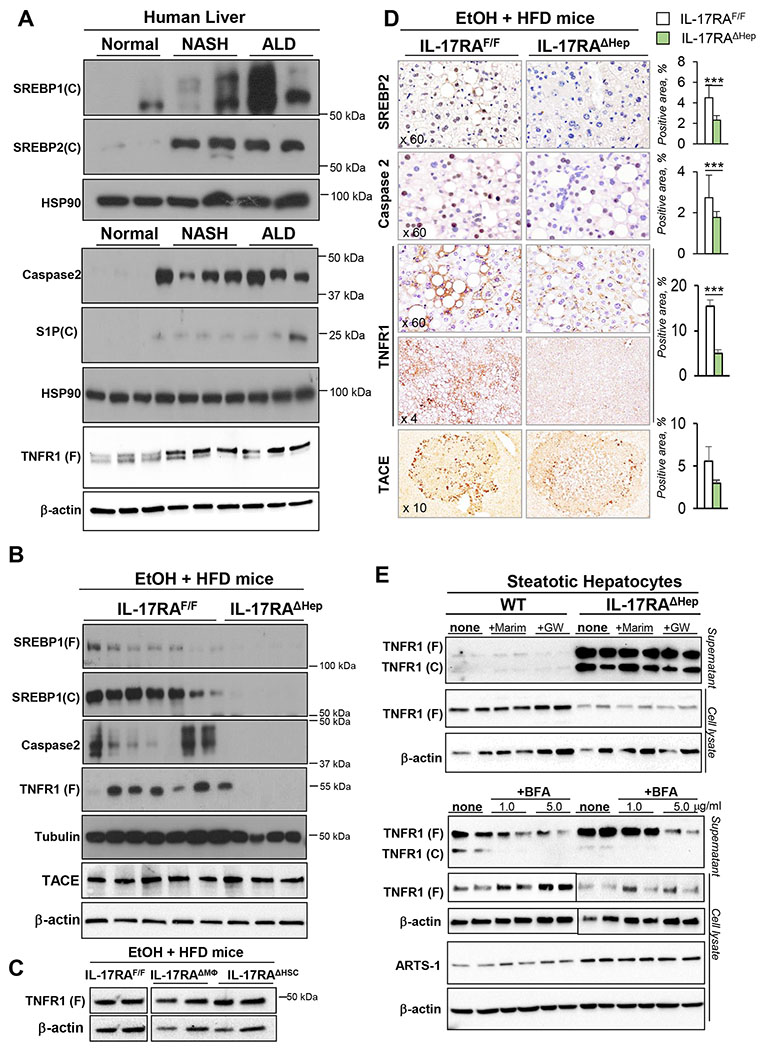
Results: We demonstrate that the global deletion of the Il-17ra gene suppressed HCC in alcohol-fed diethylnitrosamine-challenged Il-17ra-/- and major urinary protein-urokinase-type plasminogen activator/Il-17ra-/- mice compared with wild-type mice. When the cell-specific role of IL-17RA signaling was examined, the development of HCC was decreased in both alcohol-fed Il-17raΔMΦ and Il-17raΔHep mice devoid of IL-17RA in myeloid cells and hepatocytes, but not in Il-17raΔHSC mice (deficient in IL-17RA in hepatic stellate cells). Deletion of Il-17ra in myeloid cells ameliorated tumorigenesis via suppression of pro-tumorigenic/inflammatory and pro-fibrogenic responses in alcohol-fed Il-17raΔMΦ mice. Remarkably, despite a normal inflammatory response, alcohol-fed Il-17raΔHep mice developed the fewest tumors (compared with Il-17raΔMΦ mice), with reduced steatosis and fibrosis. Steatotic IL-17RA-deficient hepatocytes downregulated the expression of Cxcl1 and other chemokines, exhibited a striking defect in tumor necrosis factor (TNF)/TNF receptor 1-dependent caspase-2-SREBP1/2-DHCR7-mediated cholesterol synthesis, and upregulated the production of antioxidant vitamin D3. The pharmacological blocking of IL-17A/Th-17 cells using anti-IL-12/IL-23 antibodies suppressed the progression of HCC (by 70%) in alcohol-fed mice, indicating that targeting IL-17 signaling might provide novel strategies for the treatment of alcohol-induced HCC.
Conclusions: Overall, IL-17A is a tumor-promoting cytokine, which critically regulates alcohol-induced hepatic steatosis, inflammation, fibrosis, and HCC.
Lay summary: IL-17A is a tumor-promoting cytokine, which critically regulates inflammatory responses in macrophages (Kupffer cells and bone-marrow-derived monocytes) and cholesterol synthesis in steatotic hepatocytes in an experimental model of alcohol-induced HCC. Therefore, IL-17A may be a potential therapeutic target for patients with alcohol-induced HCC.
Keywords: ALD; Alcoholic liver disease; Cholesterol synthesis; Fibrosis; HCC; Hepatocellular carcinoma; IL-17 signaling; Inflammation; Mutational signatures.
Copyright © 2020 European Association for the Study of the Liver. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare they have no conflicts of interest. Please refer to the accompanying ICMJE disclosure forms for further details.
Figures
Comment in
-
Battling IL-17, the troublemaker in alcohol-induced hepatocellular carcinoma.J Hepatol. 2020 May;72(5):809-812. doi: 10.1016/j.jhep.2020.02.011. Epub 2020 Feb 28. J Hepatol. 2020. PMID: 32122724 No abstract available.
References
-
- Morgan RL, Baack B, Smith BD, et al. Eradication of hepatitis C virus infection and the development of hepatocellular carcinoma: a meta-analysis of observational studies. Ann Intern Med 2013;158:329–37. - PubMed
-
- Alexandrov LB. Understanding the origins of human cancer. Science 2015;350:1175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK120714/DK/NIDDK NIH HHS/United States
- R37 AI043477/AI/NIAID NIH HHS/United States
- U01 AA018663/AA/NIAAA NIH HHS/United States
- P42 ES010337/ES/NIEHS NIH HHS/United States
- R01 CA211794/CA/NCI NIH HHS/United States
- U01 AA029019/AA/NIAAA NIH HHS/United States
- R01 AA028550/AA/NIAAA NIH HHS/United States
- R56 DK088837/DK/NIDDK NIH HHS/United States
- R01 DK111866/DK/NIDDK NIH HHS/United States
- R01 DK099205/DK/NIDDK NIH HHS/United States
- U01 AA022614/AA/NIAAA NIH HHS/United States
- U01 AA027681/AA/NIAAA NIH HHS/United States
- P50 AA011999/AA/NIAAA NIH HHS/United States
- R01 AI043477/AI/NIAID NIH HHS/United States
- R01 DK101737/DK/NIDDK NIH HHS/United States
- R01 CA234128/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
