

Epigenetic Targeting of TERT-Associated Gene Expression Signature in Human Neuroblastoma with TERT Overexpression
- PMID: 31900258
- PMCID: PMC7056551
- DOI: 10.1158/0008-5472.CAN-19-2560
Epigenetic Targeting of TERT-Associated Gene Expression Signature in Human Neuroblastoma with TERT Overexpression
Abstract
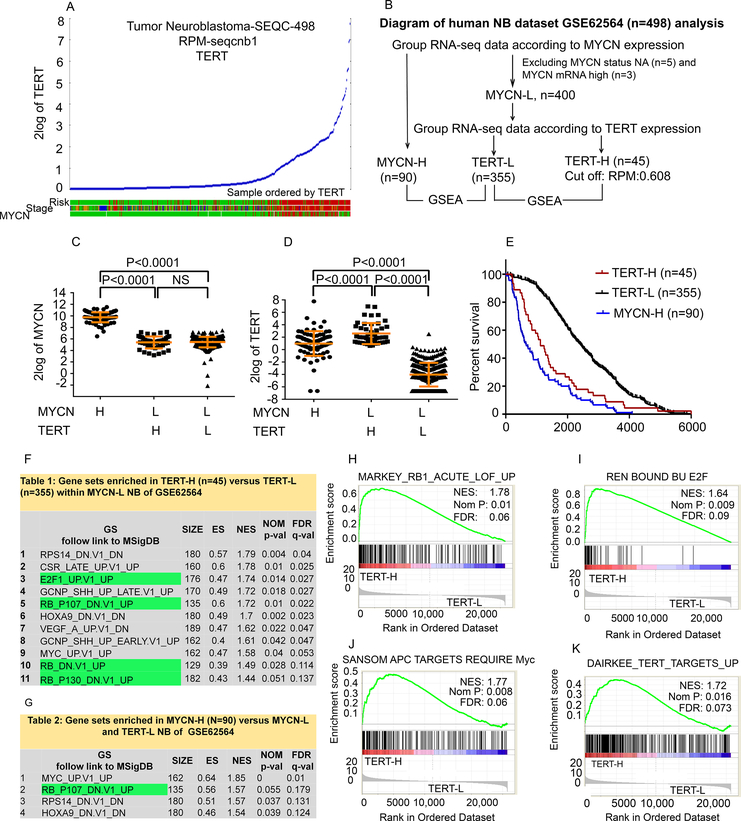
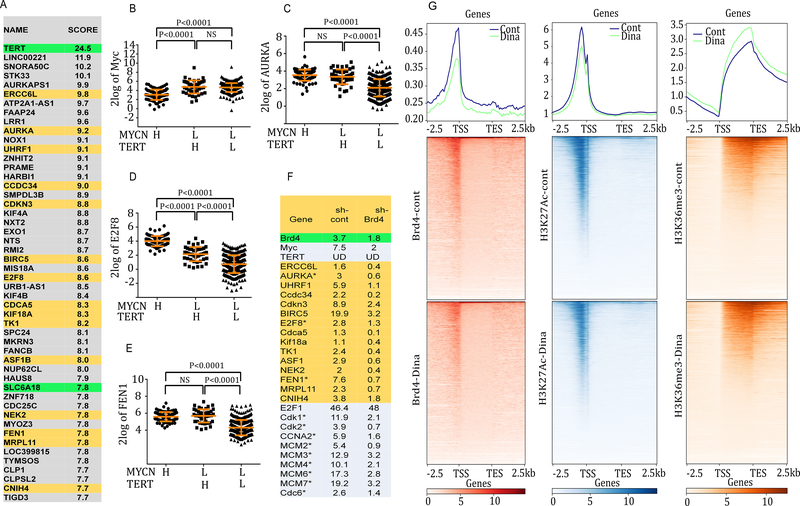
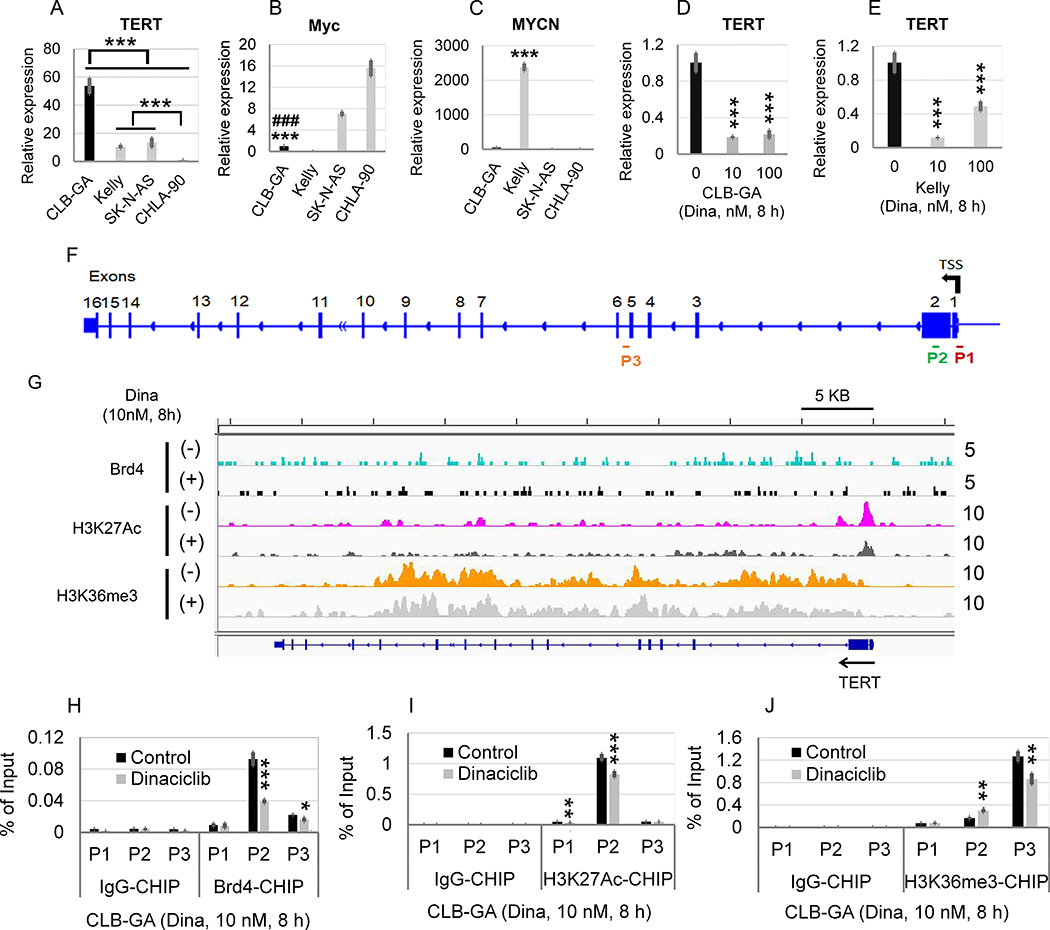
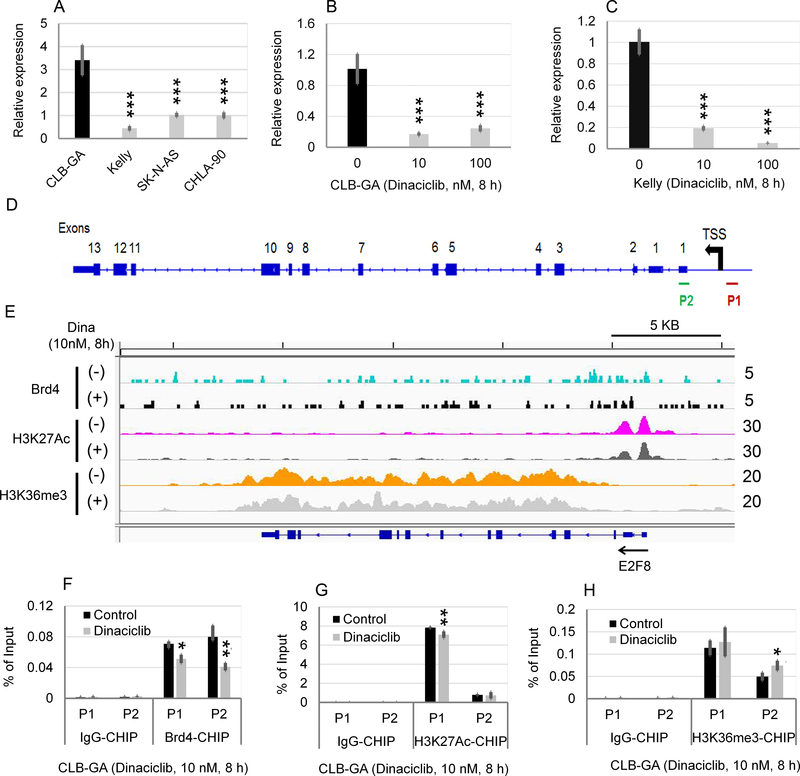
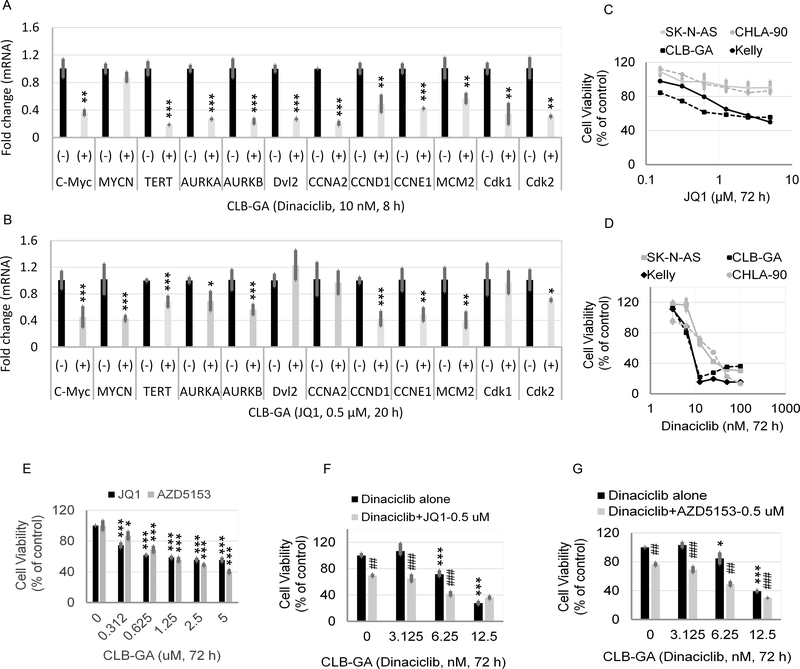
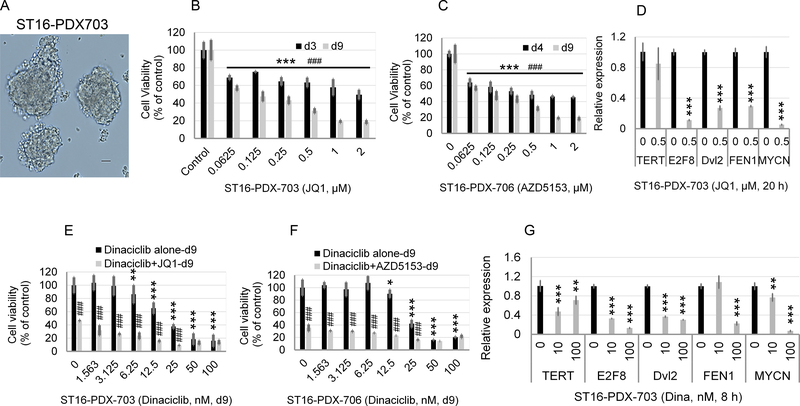
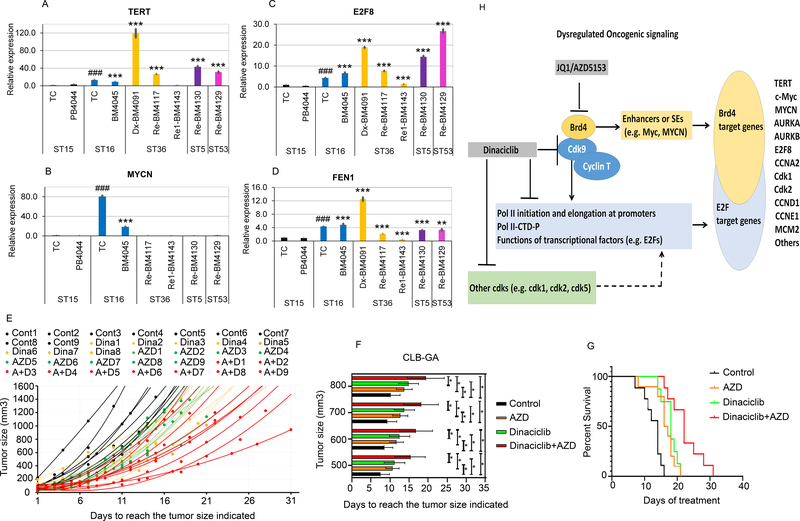
Neuroblastoma is a deadly pediatric solid tumor with infrequent recurrent somatic mutations. Particularly, the pathophysiology of tumors without MYCN amplification remains poorly defined. Utilizing an unbiased approach, we performed gene set enrichment analysis of RNA-sequencing data from 498 patients with neuroblastoma and revealed a differentially overexpressed gene signature in MYCN nonamplified neuroblastomas with telomerase reverse transcriptase (TERT) gene overexpression and coordinated activation of oncogenic signaling pathways, including E2Fs, Wnt, Myc, and the DNA repair pathway. Promoter rearrangement of the TERT gene juxtaposes the coding sequence to strong enhancer elements, leading to TERT overexpression and poor prognosis in neuroblastoma, but TERT-associated oncogenic signaling remains unclear. ChIP-seq analysis of the human CLB-GA neuroblastoma cells harboring TERT rearrangement uncovered genome-wide chromatin co-occupancy of Brd4 and H3K27Ac and robust enrichment of H3K36me3 in TERT and multiple TERT-associated genes. Brd4 and cyclin-dependent kinases (CDK) had critical regulatory roles in the expression and chromatin activation of TERT and multiple TERT-associated genes. Epigenetically targeting Brd4 or CDKs with their respective inhibitors suppressed the expression of TERT and multiple TERT-associated genes in neuroblastoma with TERT overexpression or MYCN amplification. ChIP-seq and ChIP-qPCR provided evidence that the CDK inhibitor directly inhibited Brd4 recruitment to activate chromatin globally. Therefore, inhibiting Brd4 and CDK concurrently with AZD5153 and dinaciclib would be most effective in tumor growth suppression, which we demonstrated in neuroblastoma cell lines, primary human cells, and xenografts. In summary, we describe a unique mechanism in neuroblastoma with TERT overexpression and an epigenetically targeted novel therapeutic strategy. SIGNIFICANCE: Epigenetically cotargeting Brd4 and Cdks suppresses human neuroblastoma with TERT overexpression by inhibiting the TERT-associated gene expression networks.
©2020 American Association for Cancer Research.
Conflict of interest statement
The authors declare no potential conflicts of interest.
Figures
References
-
- Matthay KK, Maris JM, Schleiermacher G, Nakagawara A, Mackall CL, Diller L, et al. Neuroblastoma. Nat Rev Dis Primers 2016, 2:16078. - PubMed
-
- Valentijn LJ, Koster J, Zwijnenburg DA, Hasselt NE, van Sluis P, Volckmann R, et al. TERT rearrangements are frequent in neuroblastoma and identify aggressive tumors. Nat Genet 2015, 47(12):1411–1414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
