

Outer Radial Glia-like Cancer Stem Cells Contribute to Heterogeneity of Glioblastoma
- PMID: 31901251
- PMCID: PMC7029801
- DOI: 10.1016/j.stem.2019.11.015
Outer Radial Glia-like Cancer Stem Cells Contribute to Heterogeneity of Glioblastoma
Abstract
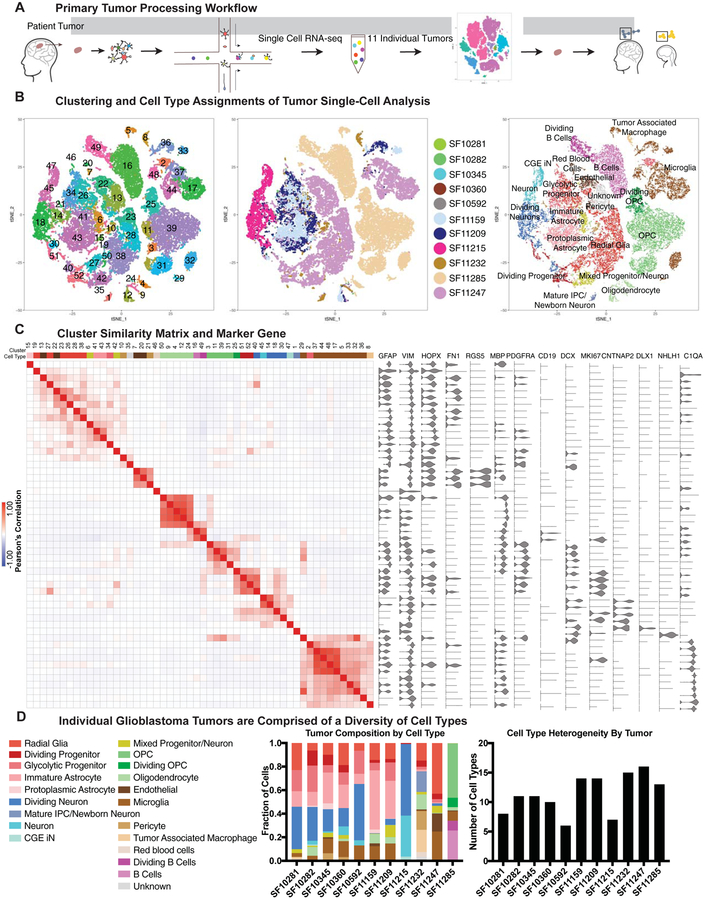
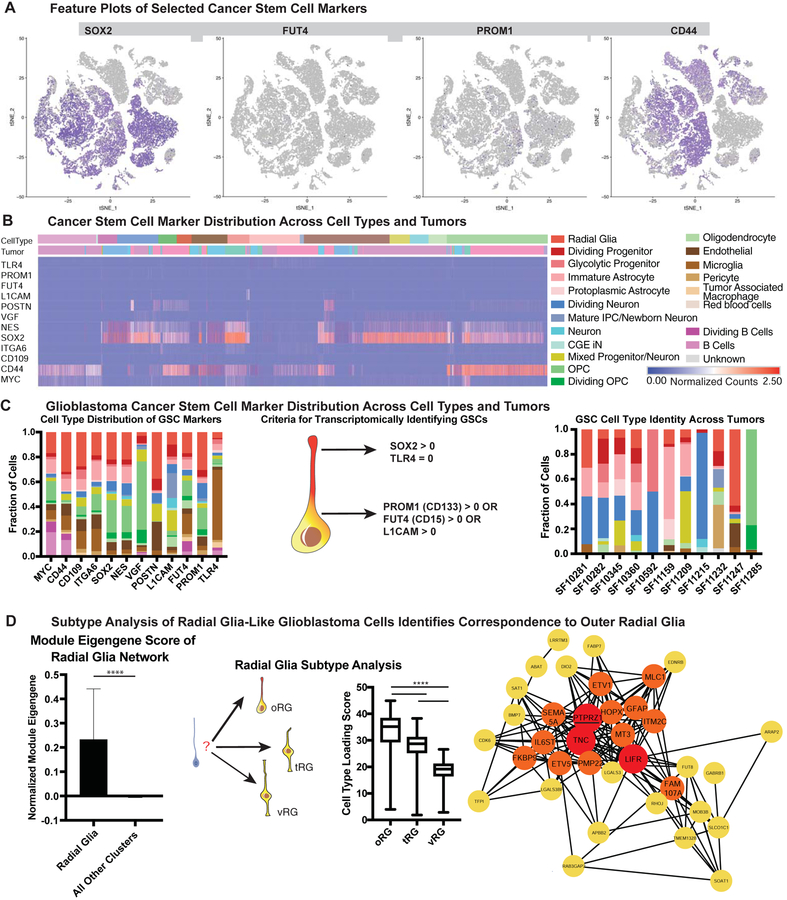
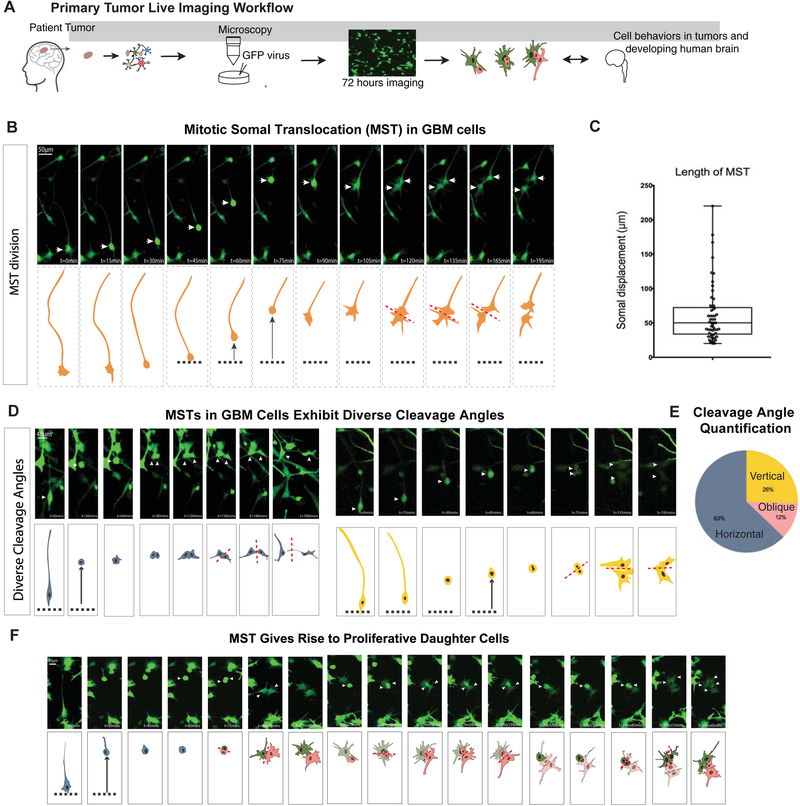
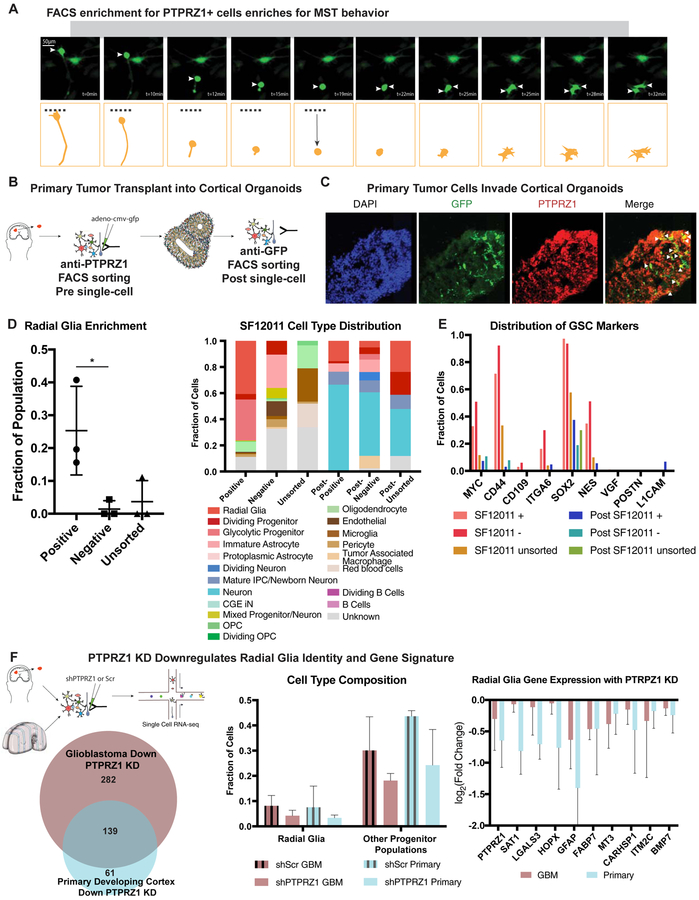
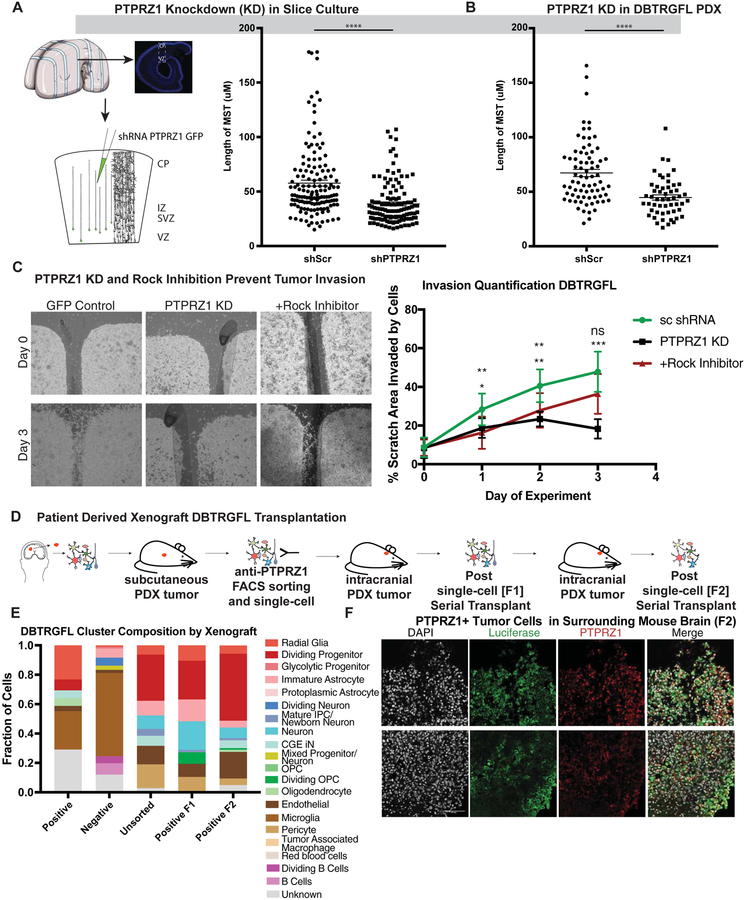
Glioblastoma is a devastating form of brain cancer. To identify aspects of tumor heterogeneity that may illuminate drivers of tumor invasion, we created a glioblastoma tumor cell atlas with single-cell transcriptomics of cancer cells mapped onto a reference framework of the developing and adult human brain. We find that multiple GSC subtypes exist within a single tumor. Within these GSCs, we identify an invasive cell population similar to outer radial glia (oRG), a fetal cell type that expands the stem cell niche in normal human cortex. Using live time-lapse imaging of primary resected tumors, we discover that tumor-derived oRG-like cells undergo characteristic mitotic somal translocation behavior previously only observed in human development, suggesting a reactivation of developmental programs. In addition, we show that PTPRZ1 mediates both mitotic somal translocation and glioblastoma tumor invasion. These data suggest that the presence of heterogeneous GSCs may underlie glioblastoma's rapid progression and invasion.
Keywords: cancer stem cell; glioblastoma; outer radial glia; single-cell sequencing; tumor heterogeneity.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures

Comment in
-
Tackling the Many Facets of Glioblastoma Heterogeneity.Cell Stem Cell. 2020 Mar 5;26(3):303-304. doi: 10.1016/j.stem.2020.02.005. Cell Stem Cell. 2020. PMID: 32142659
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
