

Mechanism of Long-Range Chromosome Motion Triggered by Gene Activation
- PMID: 31902656
- PMCID: PMC7108666
- DOI: 10.1016/j.devcel.2019.12.007
Mechanism of Long-Range Chromosome Motion Triggered by Gene Activation
Abstract
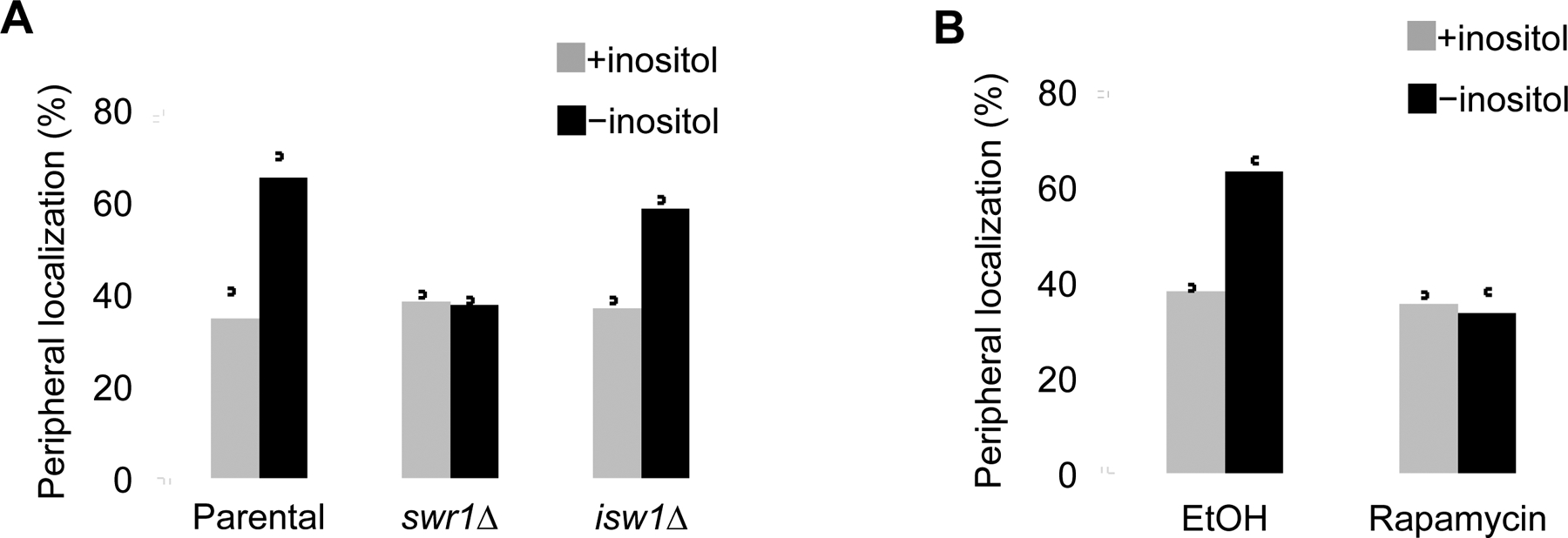
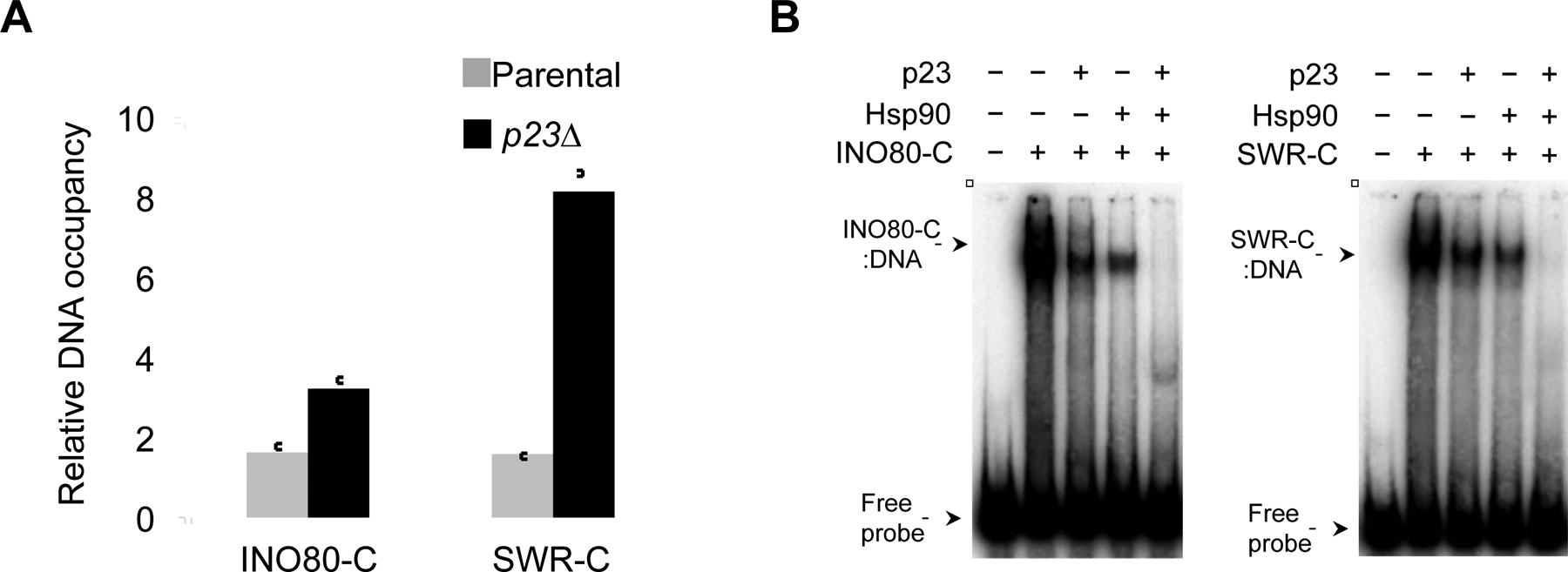
Movement of chromosome sites within interphase cells is critical for numerous pathways including RNA transcription and genome organization. Yet, a mechanism for reorganizing chromatin in response to these events had not been reported. Here, we delineate a molecular chaperone-dependent pathway for relocating activated gene loci in yeast. Our presented data support a model in which a two-authentication system mobilizes a gene promoter through a dynamic network of polymeric nuclear actin. Transcription factor-dependent nucleation of a myosin motor propels the gene locus through the actin matrix, and fidelity of the actin association was ensured by ARP-containing chromatin remodelers. Motor activity of nuclear myosin was dependent on the Hsp90 chaperone. Hsp90 further contributed by biasing the remodeler-actin interaction toward nucleosomes with the non-canonical histone H2A.Z, thereby focusing the pathway on select sites such as transcriptionally active genes. Together, the system provides a rapid and effective means to broadly yet selectively mobilize chromatin sites.
Keywords: Hsp90; chromatin motion; chromatin remodeler; genome organization; molecular chaperone; nuclear actin; nucleoskeleton.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures





Comment in
-
How Genes Move: Spatial Repositioning of Activated Genes Is Driven by Nuclear Actin-Based Pathway.Dev Cell. 2020 Feb 10;52(3):252-254. doi: 10.1016/j.devcel.2020.01.019. Dev Cell. 2020. PMID: 32049034
References
-
- Agarwal P and Zaidel-Bar R (2019). Principles of Actomyosin Regulation In Vivo. Trends in Cell Biology 29, 150–163 - PubMed
-
- Apostolou E and Thanos D (2008). Virus Infection Induces NF-κB-Dependent Interchromosomal Associations Mediating Monoallelic IFN-β Gene Expression. Cell 134(10), 85–96. - PubMed
-
- Baarlink C, Plessner M, Sherrard A, Morita K, Misu S, Virant D, Kleinschnitz E, Harniman R, Alibhai D, Baumeister S, Miyamoto K, Endesfelder U, Kaidi A and Grosse R (2017). A transient pool of nuclear F-actin at mitotic exit controls chromatin organization. Nature Cell Biology 19, 1389–1399. - PubMed
-
- Baarlink C, Wang H and Grosse R (2013). Nuclear Actin Network Assembly by Formins Regulates the SRF Coactivator MAL. Science 340, 864–867. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
