

Cuprizone-induced demyelination under physiological and post-stroke condition leads to decreased neurogenesis response in adult mouse brain
- PMID: 31904386
- PMCID: PMC9694109
- DOI: 10.1016/j.expneurol.2019.113168
Cuprizone-induced demyelination under physiological and post-stroke condition leads to decreased neurogenesis response in adult mouse brain
Abstract
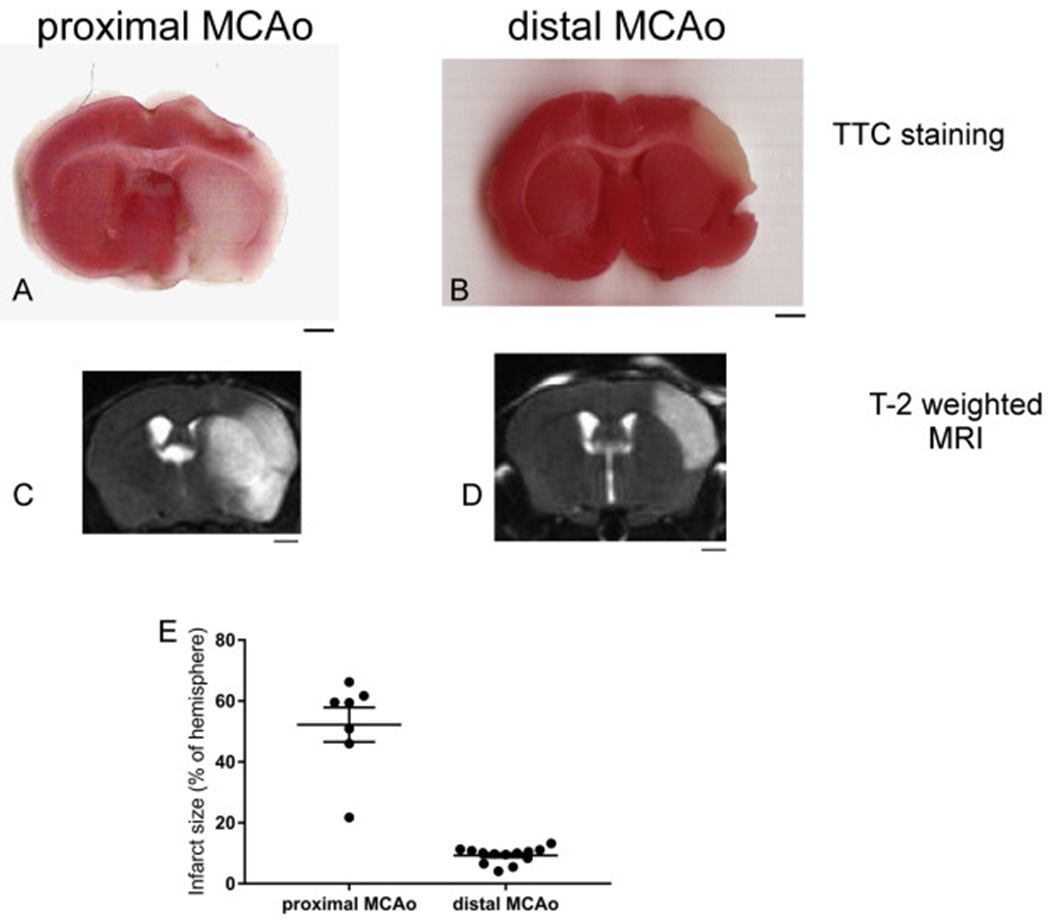
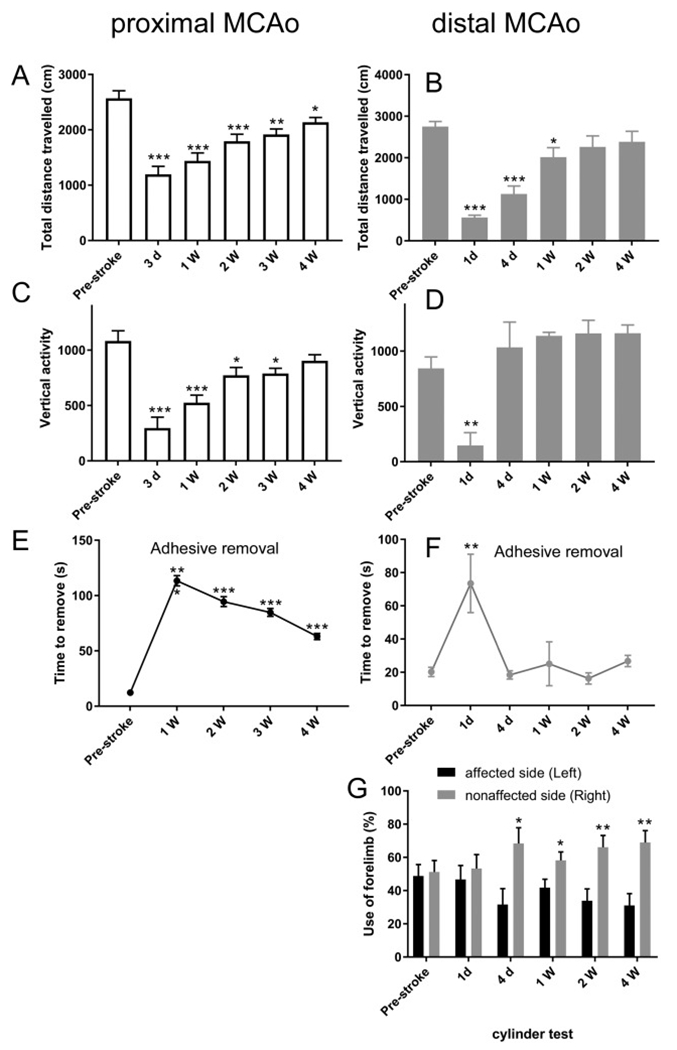
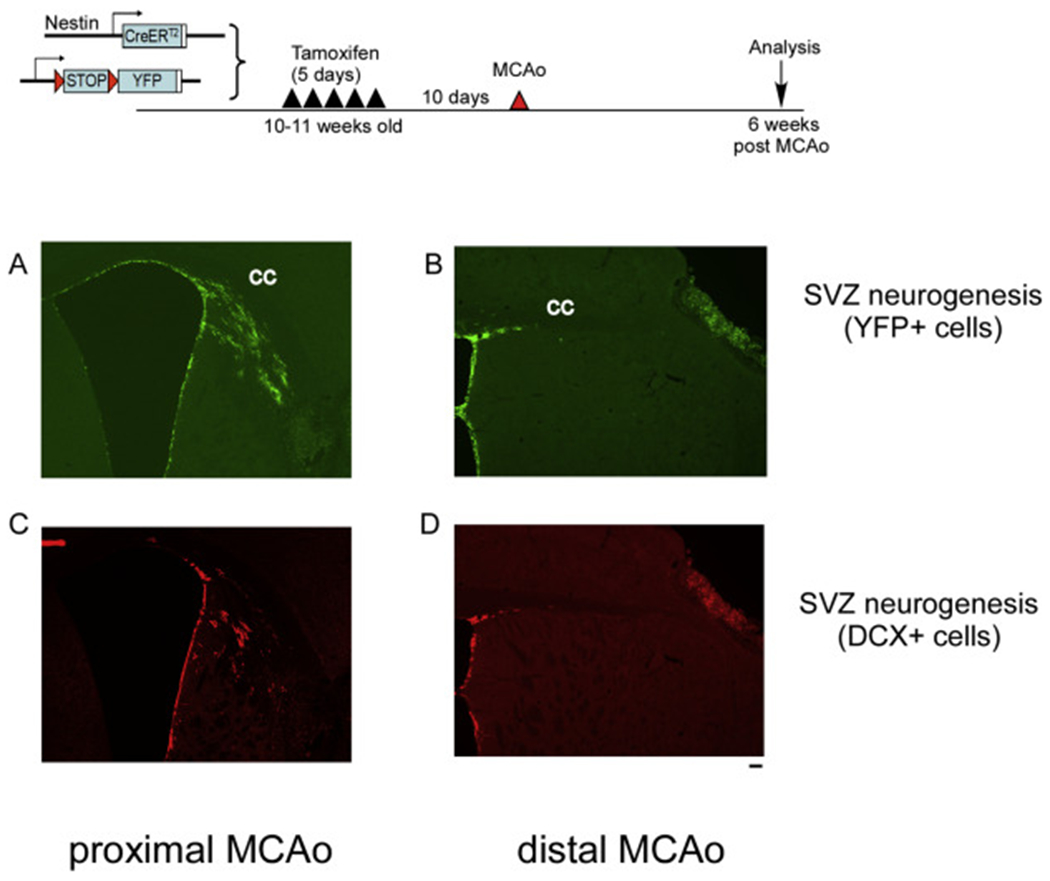
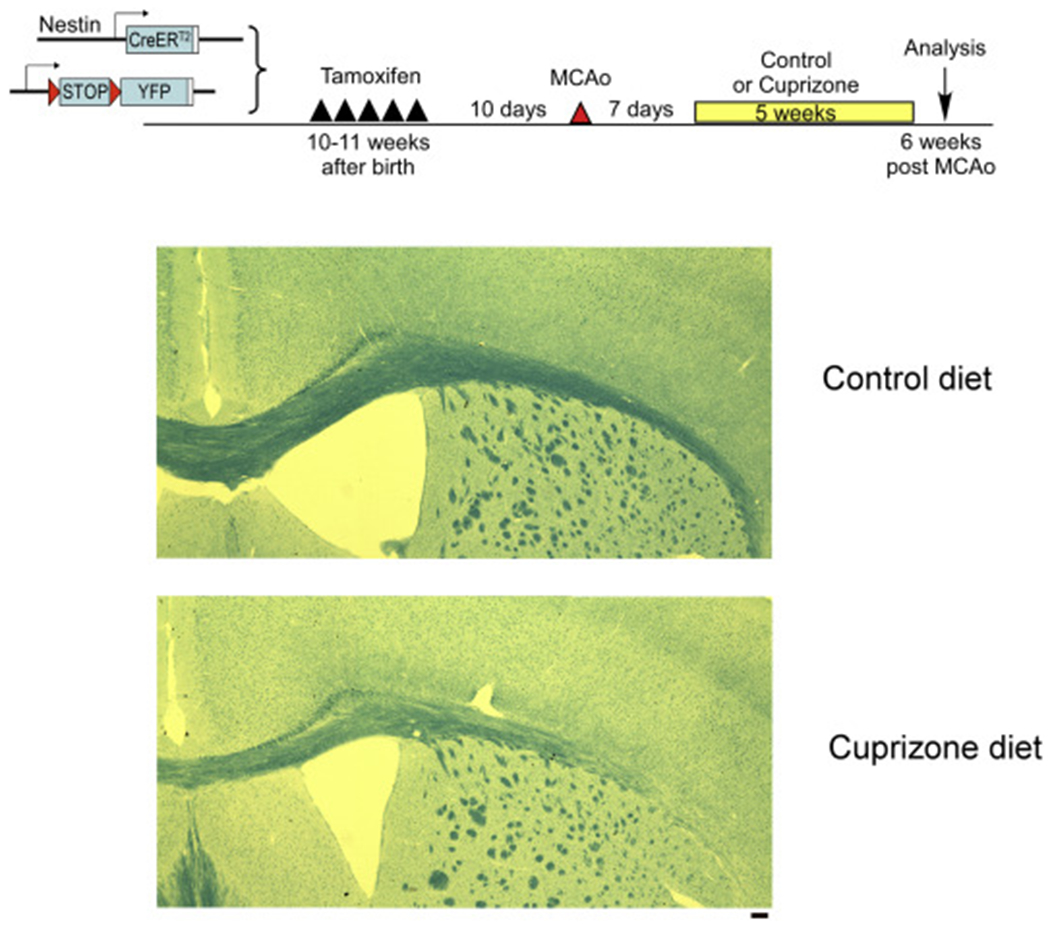
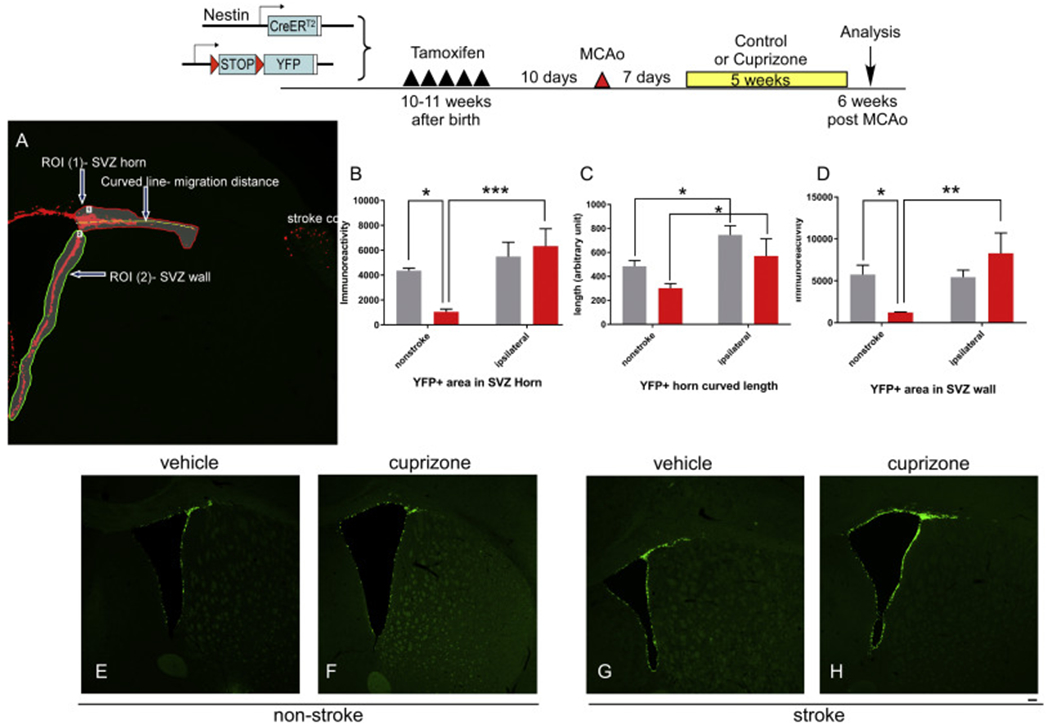
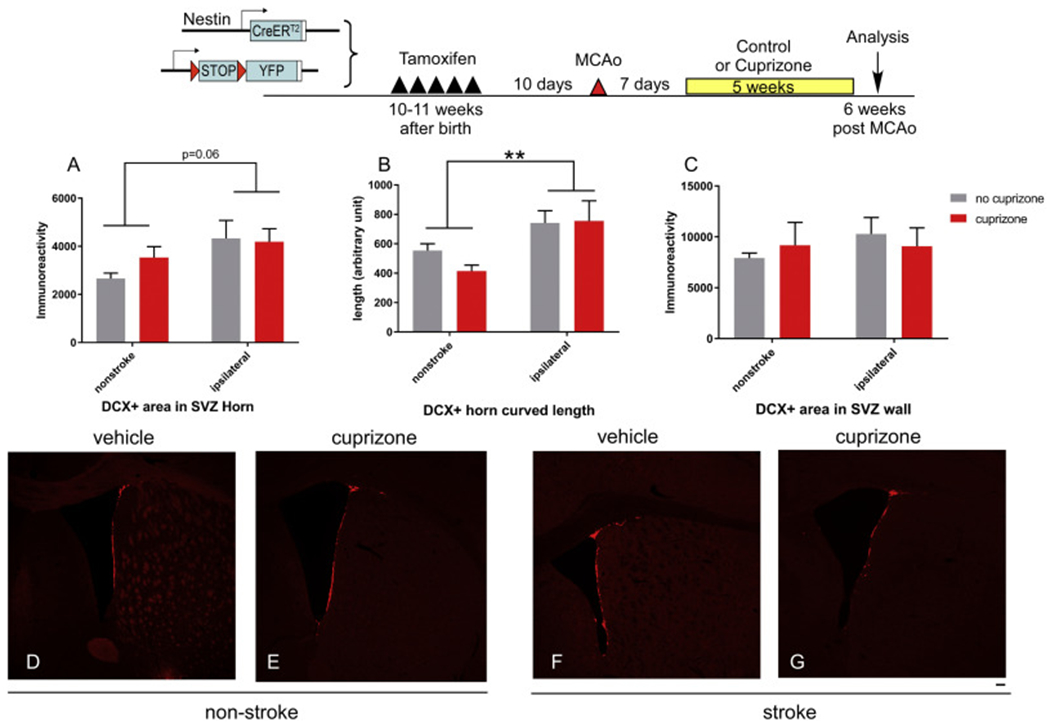
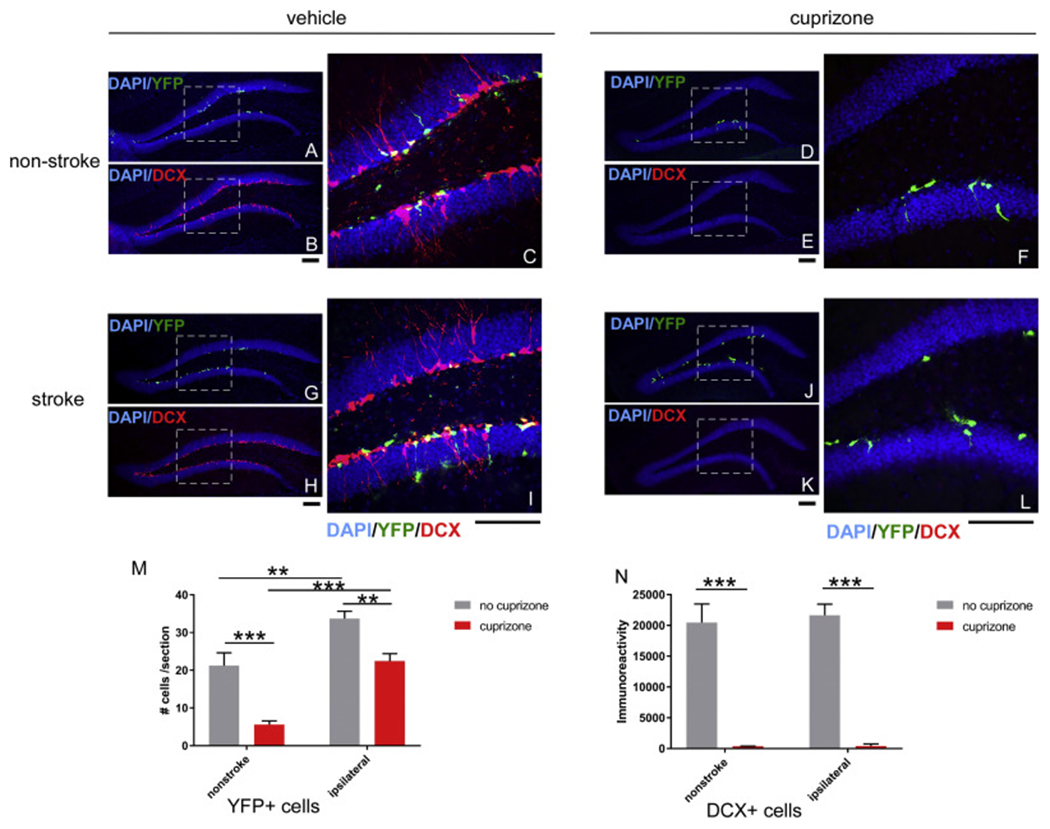
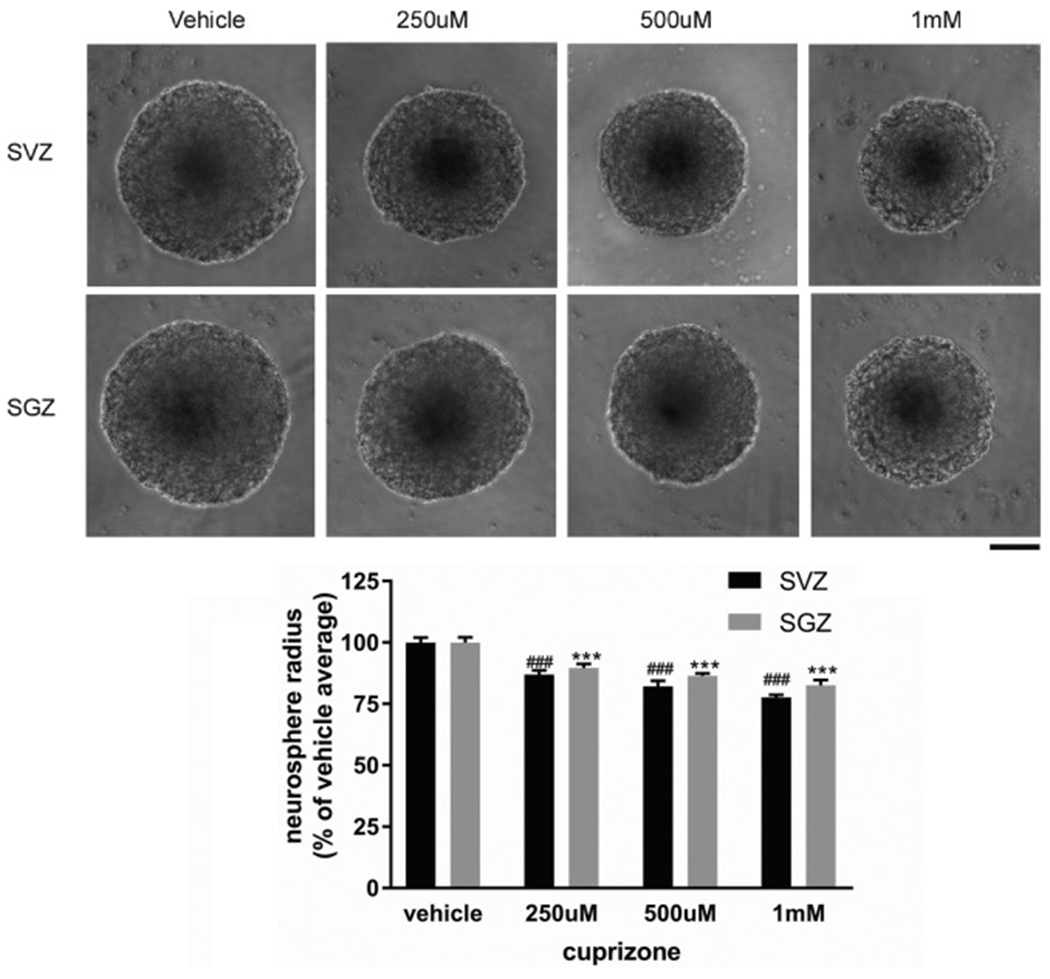
Due to the limitation in treatment window of the rtPA (recombinant tissue plasminogen activator), the development of delayed treatment for stroke is needed. We previously reported that there is a difference in neurogenesis and neuroblast migration patterns in different mouse stroke models (proximal and distal middle cerebral artery occlusion models, pMCAo or dMCAo). Specifically, compared to robust neurogenesis and substantial migration of newly born neuroblasts in pMCAo model, dMCAo only illicit limited neurogenesis and migration of neuroblasts towards ischemic area. One potential reason for this difference is the relative location of ischemic area to white matter and the neurogenic niche (subventricular zone, SVZ). Specifically, white matter could serve as a physical barrier or inhibitory factor to neurogenesis and migration in the dMCAo model. Given that a major difference in human and rodent brains is the content of white matter in the brain, in this study, we further characterize these two models and test the important hypothesis that white matter is an important contributing inhibitory factor for the limited neurogenesis in the dMCAo model. We utilized a genetically inducible NSC-specific reporter mouse line (nestin-CreERT2-R26R-YFP) to label and track NSC proliferation, survival and differentiation in ischemic brain. To test whether myelin is inhibitory to neurogenesis in dMCAo model, we demyelinated mouse brains using cuprizone treatment after stroke and examined whether there is enhanced neurogenesis or migration of neuroblasts cells in stroke mice treated with cuprizone. Our data suggests that demyelination of the brain does not result in enhanced neurogenesis or migration of neuroblasts, supporting that myelin is not a major inhibitory factor for stroke-induced neurogenesis. In addition, our results suggest that in non-stroke mice, demyelination causes decreased neurogenesis in adult brain, indicating a potential positive role of myelin in maintenance of adult neural stem cell niche.
Keywords: Distal MCAo; Neurogenesis; Proximal MCAo; Stroke; White matter.
Copyright © 2019. Published by Elsevier Inc.
Figures
References
-
- Abe H, Tanaka T, Kimura M, Mizukami S, Saito F, Imatanaka N, Akahori Y, Yoshida T, Shibutani M, 2015. Cuprizone decreases intermediate and late-stage progenitor cells in hippocampal neurogenesis of rats in a framework of 28-day oral dose toxicity study. Toxicol. Appl. Pharmacol 287 (3), 210–221. 10.1016/j.taap.2015.06.005. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
