

MicroRNAs: Biological Regulators in Pathogen-Host Interactions
- PMID: 31906500
- PMCID: PMC7016591
- DOI: 10.3390/cells9010113
MicroRNAs: Biological Regulators in Pathogen-Host Interactions
Abstract
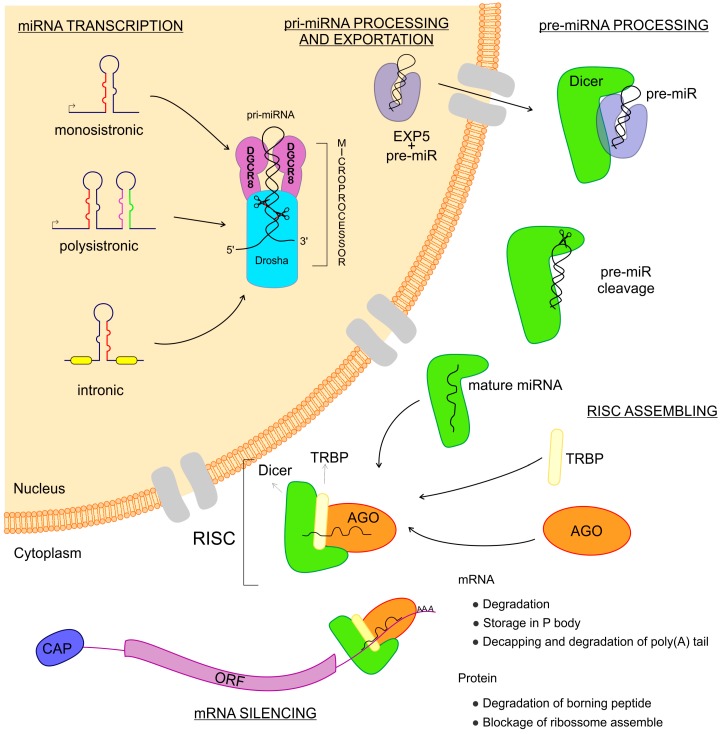
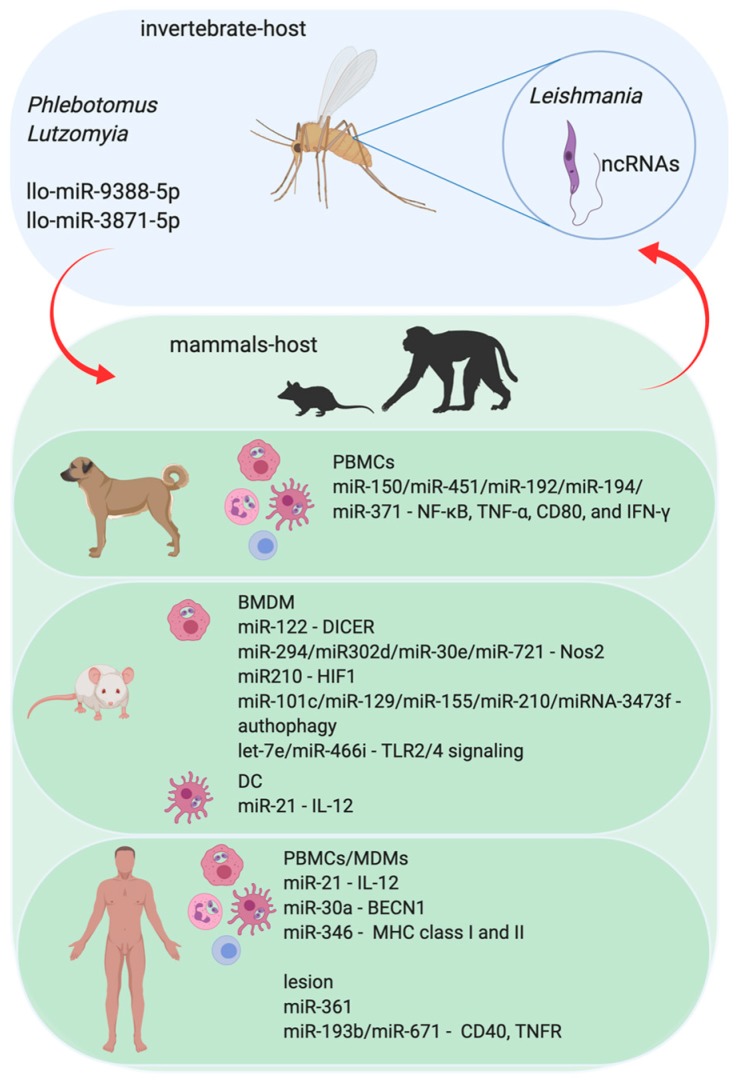
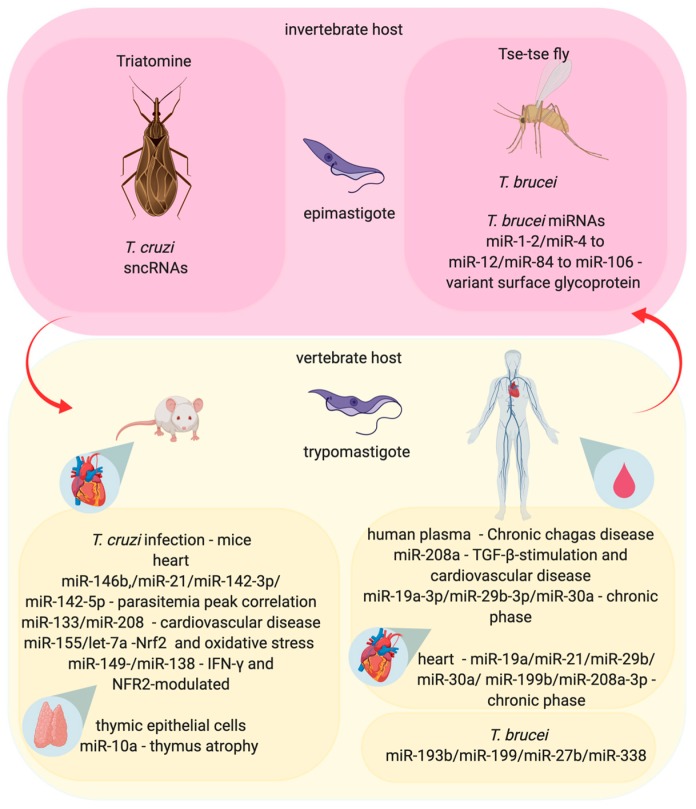
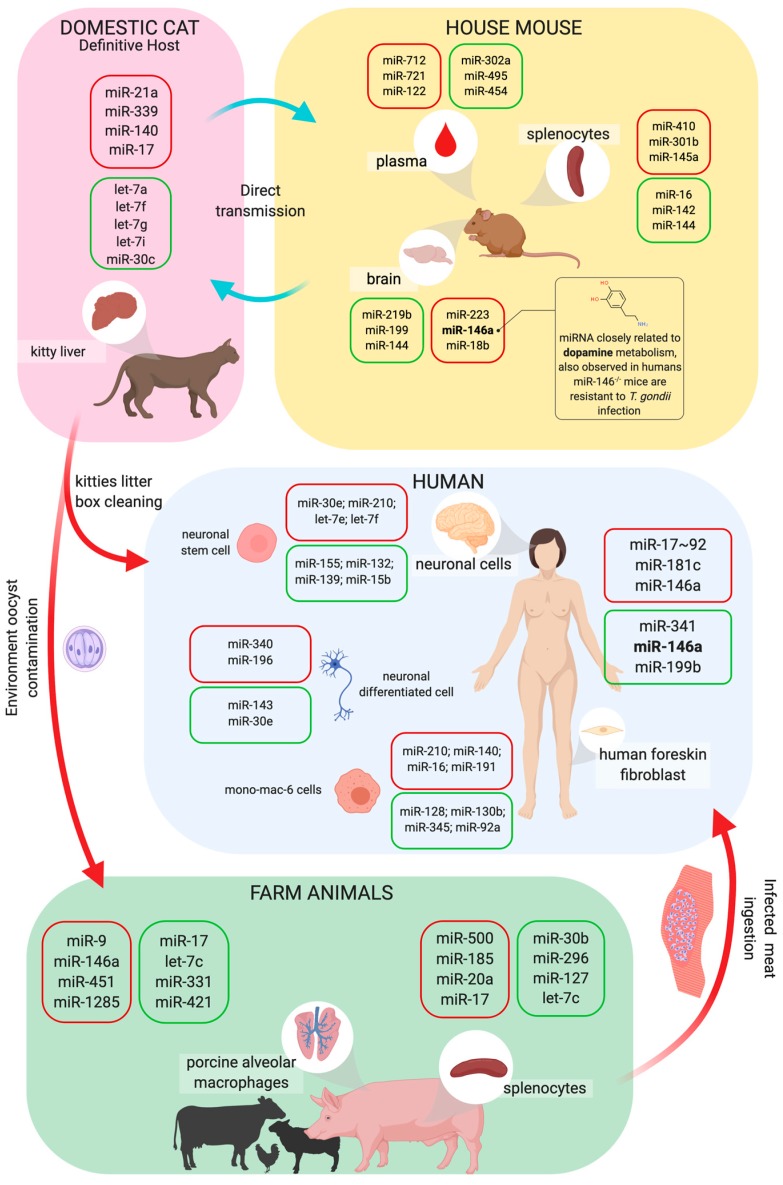
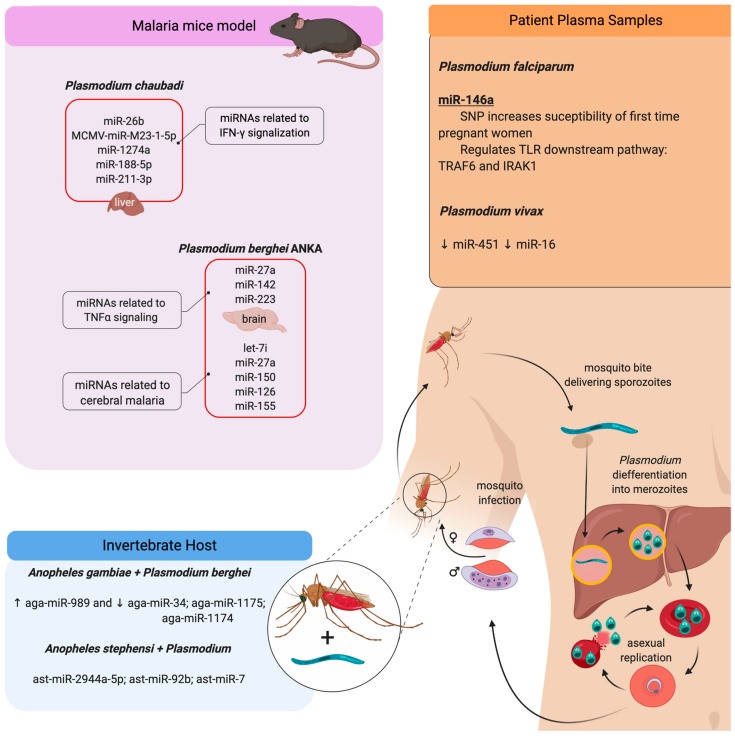
An inflammatory response is essential for combating invading pathogens. Several effector components, as well as immune cell populations, are involved in mounting an immune response, thereby destroying pathogenic organisms such as bacteria, fungi, viruses, and parasites. In the past decade, microRNAs (miRNAs), a group of noncoding small RNAs, have emerged as functionally significant regulatory molecules with the significant capability of fine-tuning biological processes. The important role of miRNAs in inflammation and immune responses is highlighted by studies in which the regulation of miRNAs in the host was shown to be related to infectious diseases and associated with the eradication or susceptibility of the infection. Here, we review the biological aspects of microRNAs, focusing on their roles as regulators of gene expression during pathogen-host interactions and their implications in the immune response against Leishmania, Trypanosoma, Toxoplasma, and Plasmodium infectious diseases.
Keywords: gene expression; host; immune response; microRNAs; parasite; pathogen; post-transcriptional.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Derrien T., Johnson R., Bussotti G., Tanzer A., Djebali S., Tilgner H., Guernec G., Martin D., Merkel A., Knowles D.G., et al. The gencode v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012;22:1775–1789. doi: 10.1101/gr.132159.111. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
