

Specialized stellate cells offer a privileged route for rapid water flux in Drosophila renal tubule
- PMID: 31907321
- PMCID: PMC6983416
- DOI: 10.1073/pnas.1915943117
Specialized stellate cells offer a privileged route for rapid water flux in Drosophila renal tubule
Abstract
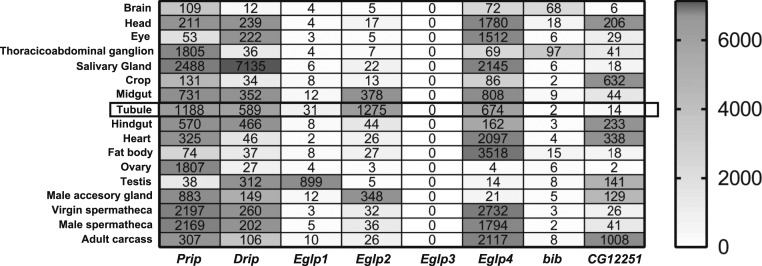
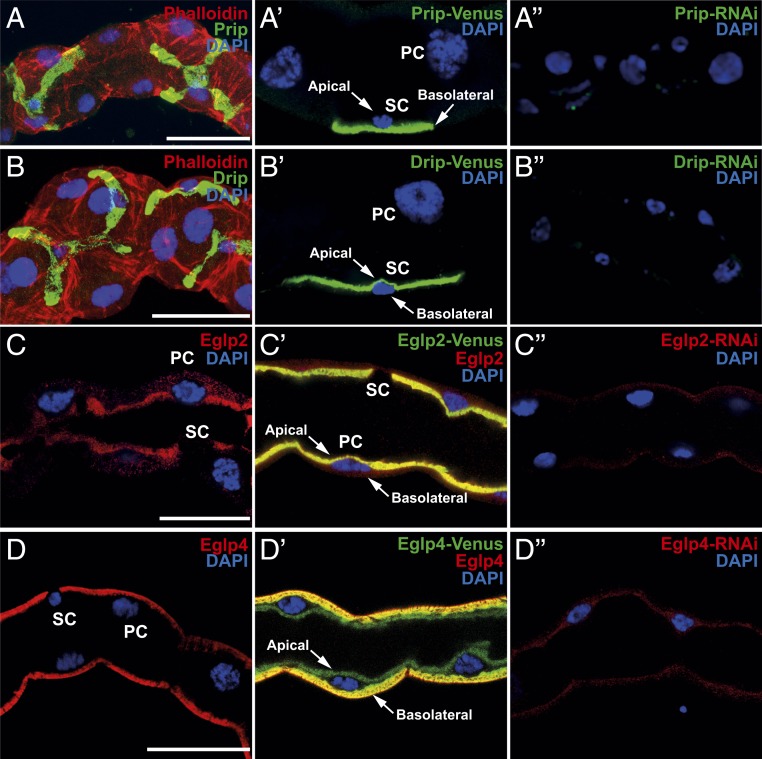
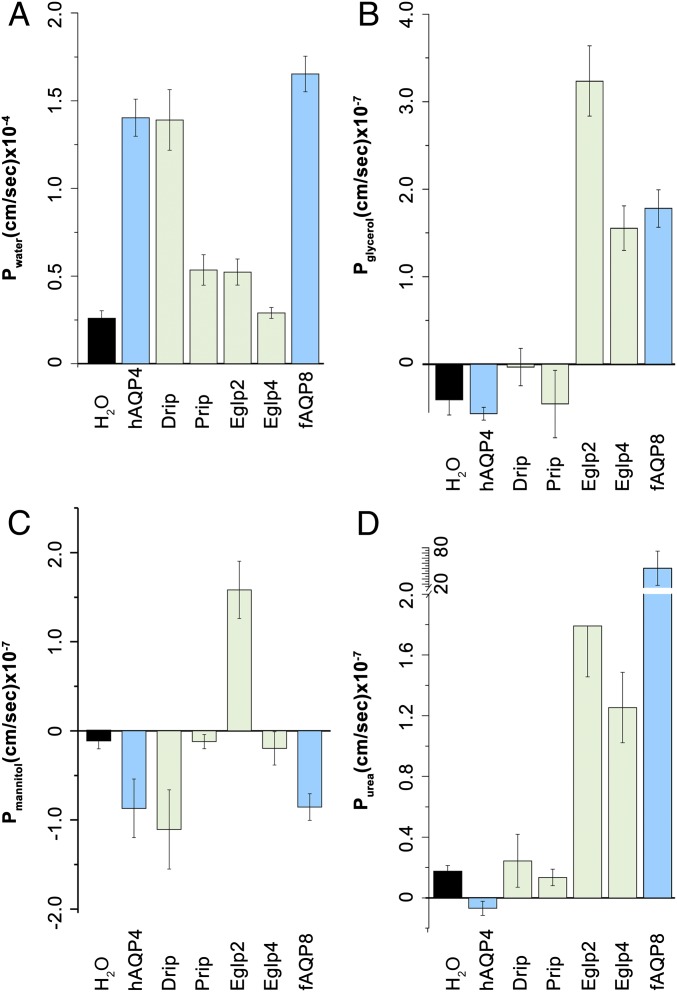
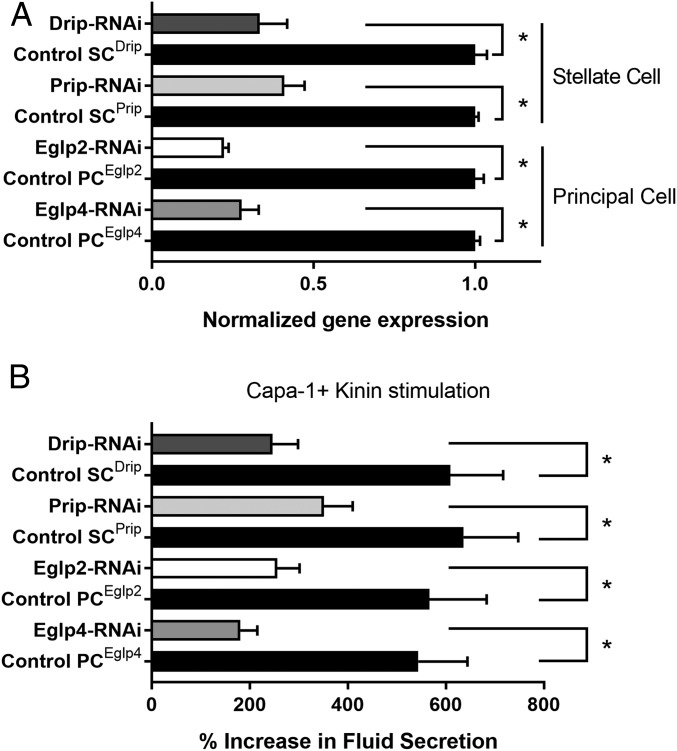
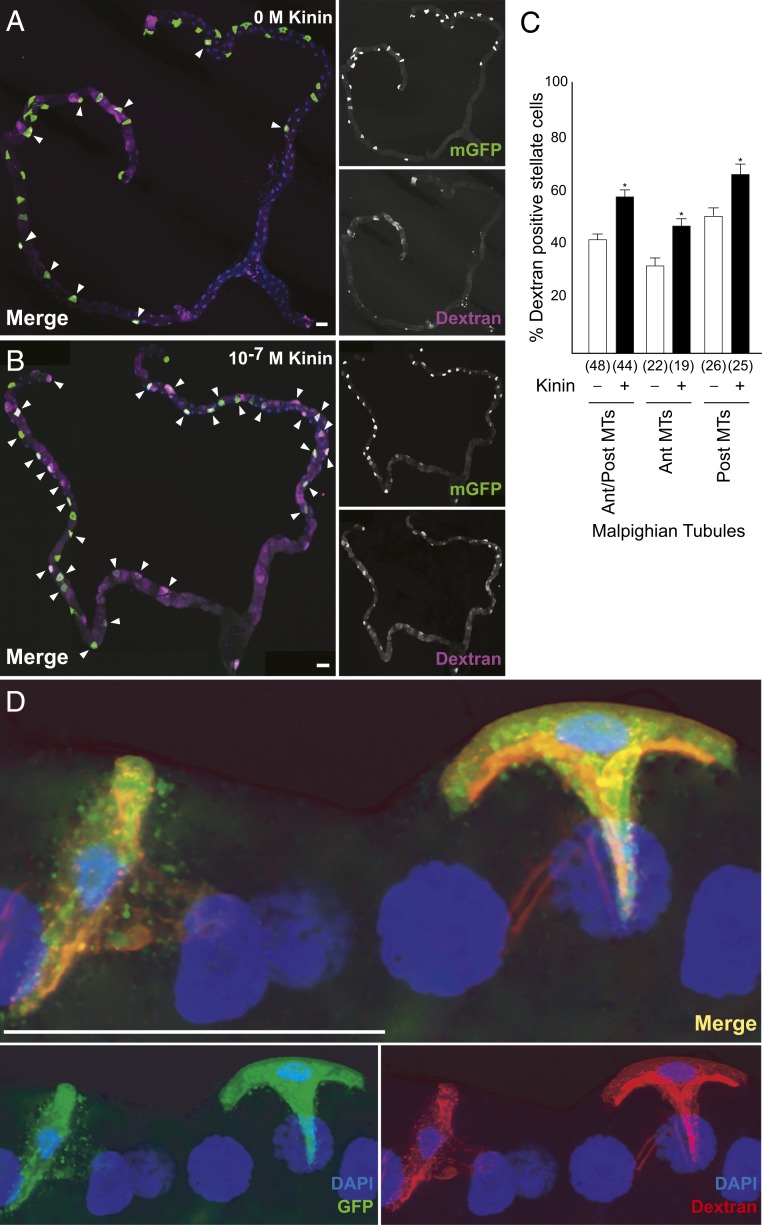
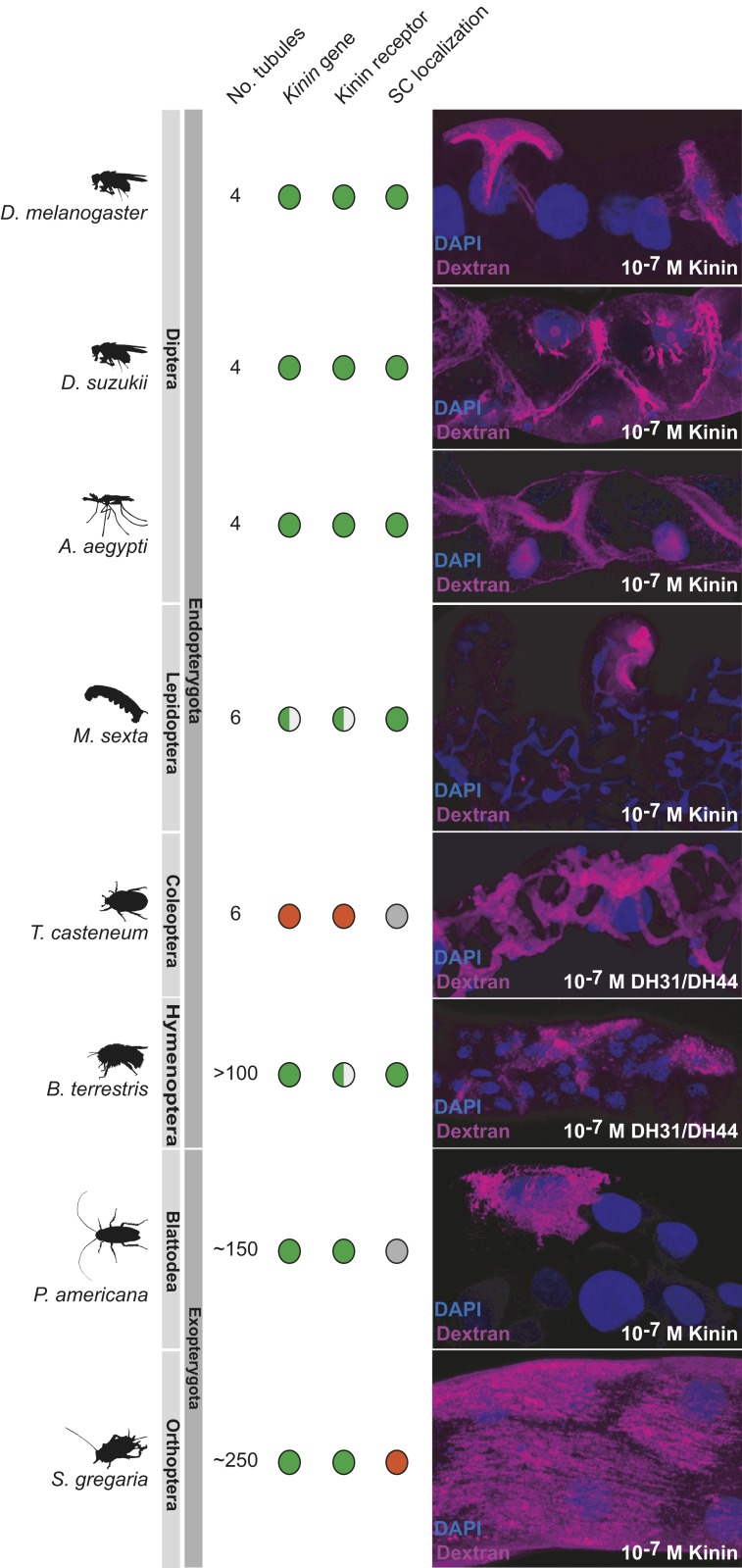
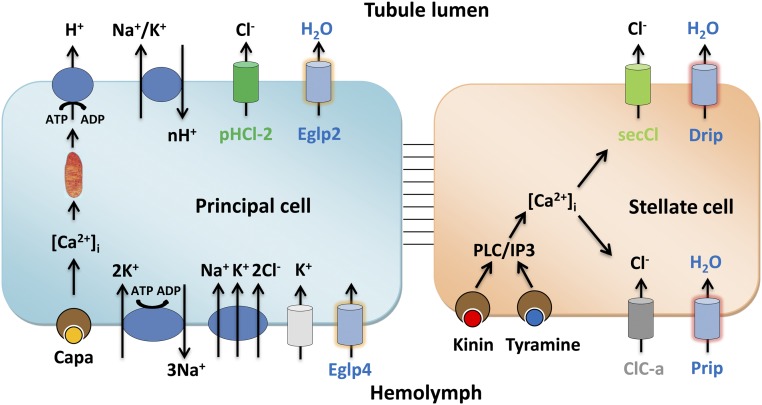
Insects are highly successful, in part through an excellent ability to osmoregulate. The renal (Malpighian) tubules can secrete fluid faster on a per-cell basis than any other epithelium, but the route for these remarkable water fluxes has not been established. In Drosophila melanogaster, we show that 4 genes of the major intrinsic protein family are expressed at a very high level in the fly renal tissue: the aquaporins (AQPs) Drip and Prip and the aquaglyceroporins Eglp2 and Eglp4 As predicted from their structure, and by their transport function by expressing these proteins in Xenopus oocytes, Drip, Prip, and Eglp2 show significant and specific water permeability, whereas Eglp2 and Eglp4 show very high permeability to glycerol and urea. Knockdowns of any of these genes result in impaired hormone-induced fluid secretion. The Drosophila tubule has 2 main secretory cell types: active cation-transporting principal cells, wherein the aquaglyceroporins localize to opposite plasma membranes, and small stellate cells, the site of the chloride shunt conductance, with these AQPs localizing to opposite plasma membranes. This suggests a model in which osmotically obliged water flows through the stellate cells. Consistent with this model, fluorescently labeled dextran, an in vivo marker of membrane water permeability, is trapped in the basal infoldings of the stellate cells after kinin diuretic peptide stimulation, confirming that these cells provide the major route for transepithelial water flux. The spatial segregation of these components of epithelial water transport may help to explain the unique success of the higher insects in regulating their internal environments.
Keywords: Drosophila melanogaster; Malpighian tubule; Xenopus oocyte; aquaporin; stellate cell.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Maddrell S. H. P., The fastest fluid-secreting cell known: The upper Malpighian tubule cell of Rhodnius. BioEssays 13, 357–362 (1991).
-
- Dow J. A. T., et al. , The Malpighian tubules of Drosophila melanogaster: A novel phenotype for studies of fluid secretion and its control. J. Exp. Biol. 197, 421–428 (1994). - PubMed
-
- O’Donnell M. J., Aldis G. K., Maddrell S. H. P., Measurements of osmotic permeability in the Malpighian tubules of an insect, Rhodnius prolixus Stal. Proc. R. Soc. Lond. B Biol. Sci. 216, 267–277 (1982).
-
- Beyenbach K. W., Skaer H., Dow J. A. T., The developmental, molecular, and transport biology of Malpighian tubules. Annu. Rev. Entomol. 55, 351–374 (2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/P008097/1 /BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/L002647/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- R25 DK101405/DK/NIDDK NIH HHS/United States
- MC_PC_17190/MRC_/Medical Research Council/United Kingdom
- U54 DK100227/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
