

Lipid droplet motility and organelle contacts
- PMID: 31909374
- PMCID: PMC6943980
- DOI: 10.1177/2515256419895688
Lipid droplet motility and organelle contacts
Abstract
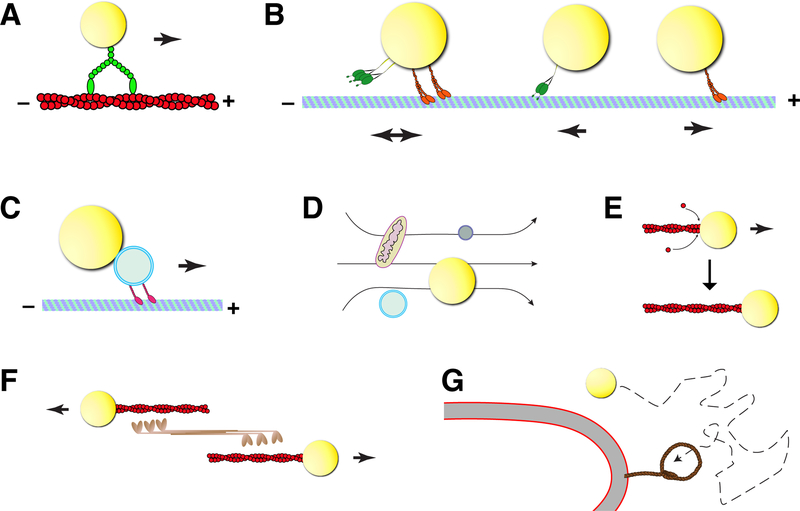
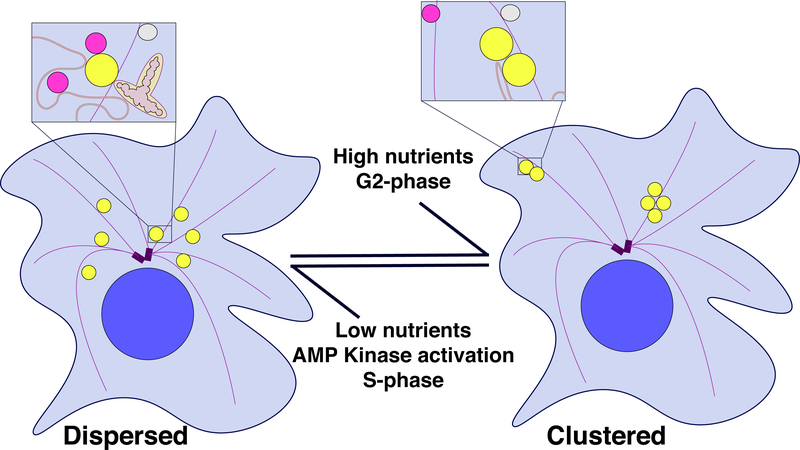
Lipid droplets (LDs) are fat storage organelles integral to energy homeostasis and a wide range of cellular processes. LDs physically and functionally interact with many partner organelles, including the ER, mitochondria, lysosomes, and peroxisomes. Recent findings suggest that the dynamics of LD inter-organelle contacts is in part controlled by LD intracellular motility. LDs can be transported directly by motor proteins along either actin filaments or microtubules, via Kinesin-1, Cytoplasmic Dynein, and type V Myosins. LDs can also be propelled indirectly, by hitchhiking on other organelles, cytoplasmic flows, and potentially actin polymerization. Although the anchors that attach motors to LDs remain elusive, other regulators of LD motility have been identified, ranging from modification of the tracks to motor co-factors to members of the perilipin family of LD proteins. Manipulating these regulatory pathways provides a tool to probe whether altered motility affects organelle contacts and has revealed that LD motility can promote interactions with numerous partners, with profound consequences for metabolism. LD motility can cause dramatic redistribution of LDs between a clustered and a dispersed state, resulting in altered organelle contacts and LD turnover. We propose that LD motility can thus promote switches in the metabolic state of a cell. Finally, LD motility is also important for LD allocation during cell division. In a number of animal embryos, uneven allocation results in a large difference in LD content in distinct daughter cells, suggesting cell-type specific LD needs.
Keywords: cell division; contact; cytoskeleton; lipid droplet; metabolism; molecular motor.
Figures
References
-
- Akil A, Peng J, Omrane M, Gondeau C, Desterke C, Marin M, Tronchere H, Taveneau C, Sar S, Briolotti P, et al. (2016). Septin 9 induces lipid droplets growth by a phosphatidylinositol-5-phosphate and microtubule-dependent mechanism hijacked by HCV. Nat Commun 7, 12203. doi: 10.1038/ncomms12203 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials