

An intact keratin network is crucial for mechanical integrity and barrier function in keratinocyte cell sheets
- PMID: 31912195
- PMCID: PMC11104923
- DOI: 10.1007/s00018-019-03424-7
An intact keratin network is crucial for mechanical integrity and barrier function in keratinocyte cell sheets
Abstract
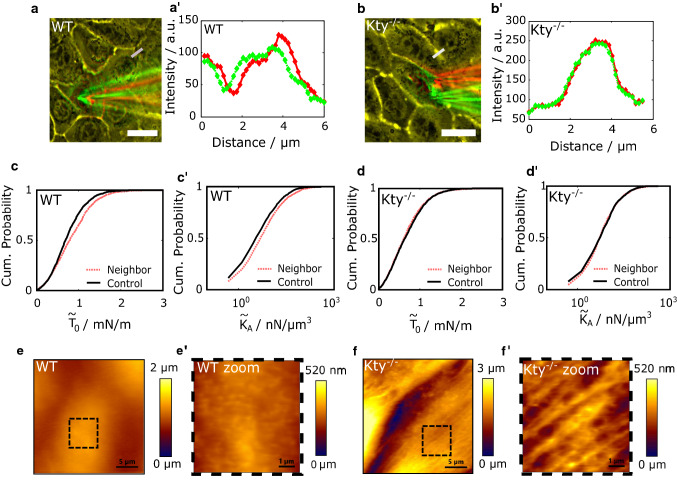
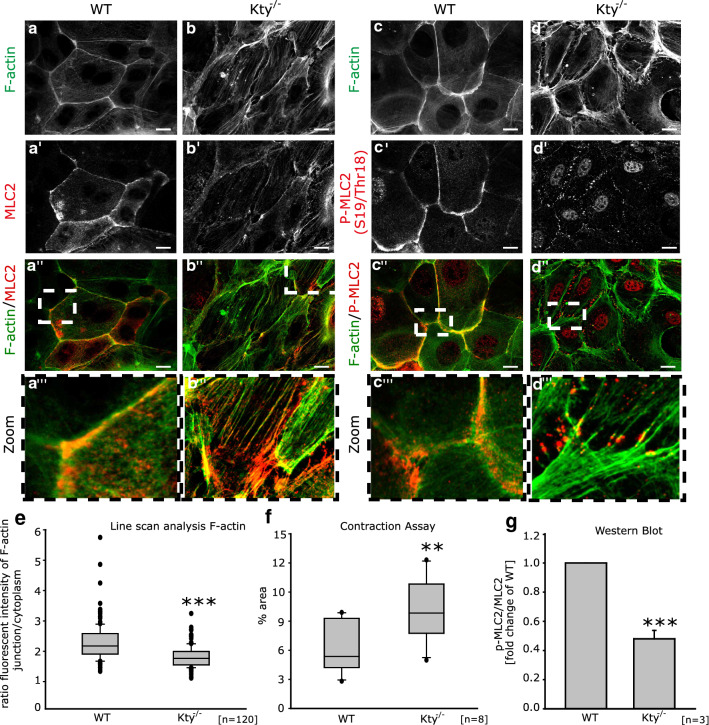
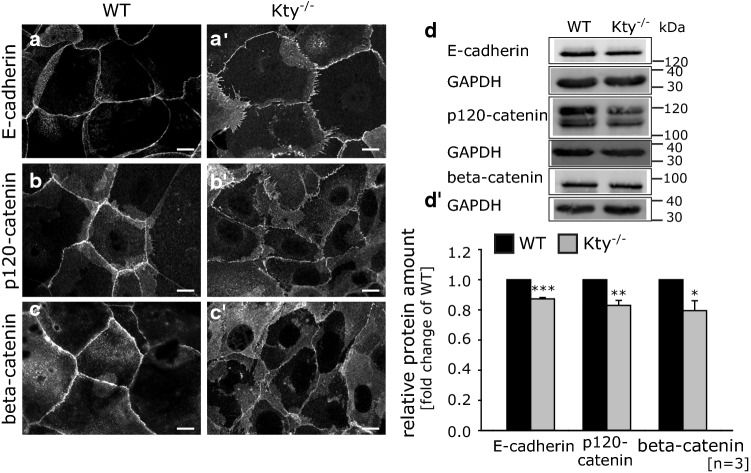
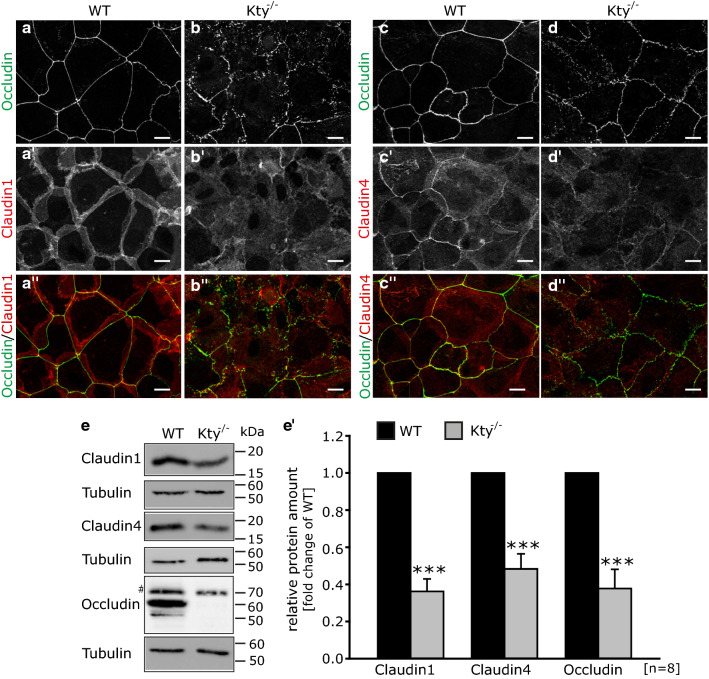
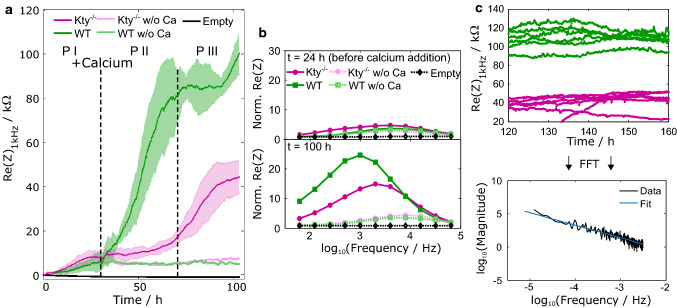
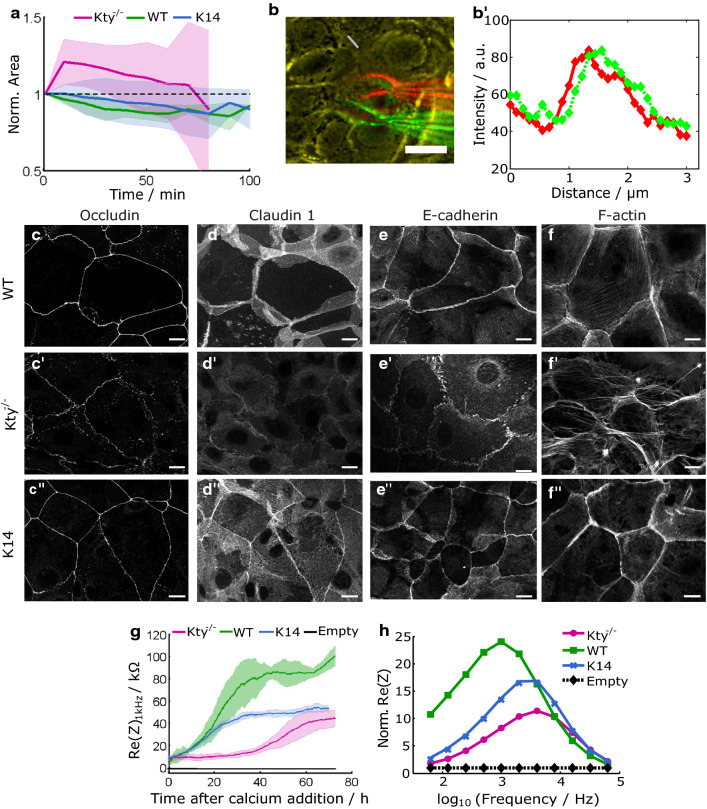
The isotype-specific composition of the keratin cytoskeleton is important for strong adhesion, force resilience, and barrier function of the epidermis. However, the mechanisms by which keratins regulate these functions are still incompletely understood. In this study, the role and significance of the keratin network for mechanical integrity, force transmission, and barrier formation were analyzed in murine keratinocytes. Following the time-course of single-cell wound closure, wild-type (WT) cells slowly closed the gap in a collective fashion involving tightly connected neighboring cells. In contrast, the mechanical response of neighboring cells was compromised in keratin-deficient cells, causing an increased wound area initially and an inefficient overall wound closure. Furthermore, the loss of the keratin network led to impaired, fragmented cell-cell junctions, and triggered a profound change in the overall cellular actomyosin architecture. Electric cell-substrate impedance sensing of cell junctions revealed a dysfunctional barrier in knockout (Kty-/-) cells compared to WT cells. These findings demonstrate that Kty-/- cells display a novel phenotype characterized by loss of mechanocoupling and failure to form a functional barrier. Re-expression of K5/K14 rescued the barrier defect to a significant extent and reestablished the mechanocoupling with remaining discrepancies likely due to the low abundance of keratins in that setting. Our study reveals the major role of the keratin network for mechanical homeostasis and barrier functionality in keratinocyte layers.
Keywords: Barrier functionality; Keratin cytoskeleton; Mechanical homeostasis; Wound closure.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
