

An RXLR effector PlAvh142 from Peronophythora litchii triggers plant cell death and contributes to virulence
- PMID: 31912634
- PMCID: PMC7036370
- DOI: 10.1111/mpp.12905
An RXLR effector PlAvh142 from Peronophythora litchii triggers plant cell death and contributes to virulence
Abstract
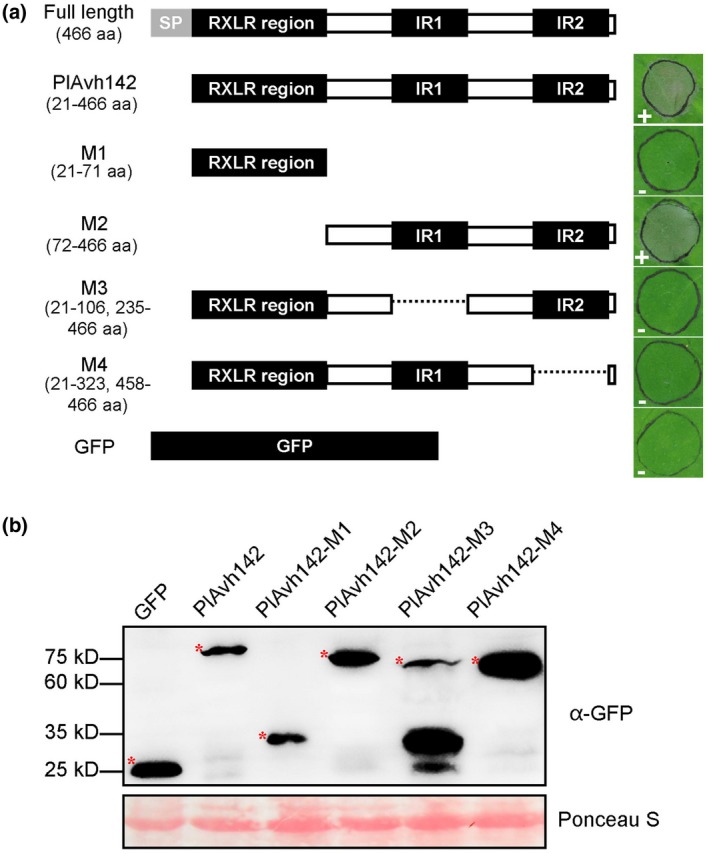
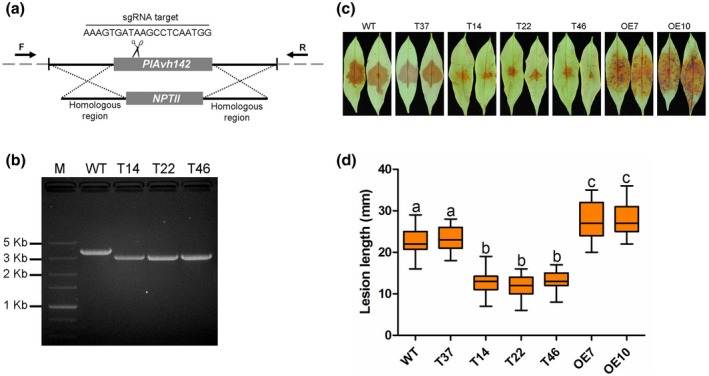
Litchi downy blight, caused by the phytopathogenic oomycete Peronophythora litchii, results in tremendous economic loss in litchi production every year. To successfully colonize the host cell, Phytophthora species secret hundreds of RXLR effectors that interfere with plant immunity and facilitate the infection process. Previous work has already predicted 245 candidate RXLR effector-encoding genes in P. litchii, 212 of which have been cloned and tested for plant cell death-inducing activity in this study. We found three such RXLR effectors could trigger plant cell death through transient expression in Nicotiana benthamiana. Further experiments demonstrated that PlAvh142 could induce cell death and immune responses in several plants. We also found that PlAvh142 localized in both the cytoplasm and nucleus of plant cells. The cytoplasmic localization was critical for its cell death-inducing activity. Moreover, deletion either of the two internal repeats in PlAvh142 abolished the cell death-inducing activity. Virus-induced gene silencing assays showed that cell death triggered by PlAvh142 was dependent on the plant transduction components RAR1 (require for Mla12 resistance), SGT1 (suppressor of the G2 allele of skp1) and HSP90 (heat shock protein 90). Finally, knockout of PlAvh142 resulted in significantly attenuated P. litchii virulence on litchi plants, whereas the PlAvh142-overexpressed mutants were more aggressive. These data indicated that PlAvh142 could be recognized in plant cytoplasm and is an important virulence RXLR effector of P. litchii.
Keywords: Peronophythora litchii; RXLR effector; cell death; plant immunity.
© 2020 The Authors. Molecular Plant Pathology published by British Society for Plant Pathology and John Wiley & Sons Ltd.
Figures





References
-
- Andrade, M.A. , Perez‐Iratxeta, C. and Ponting, C.P. (2001) Protein repeats: structures, functions, and evolution. Journal of Structural Biology, 134, 117–131. - PubMed
-
- Asai, S. and Yoshioka, H. (2009) Nitric oxide as a partner of reactive oxygen species participates in disease resistance to necrotrophic pathogen Botrytis cinerea in Nicotiana benthamiana . Molecular Plant‐Microbe Interactions, 22, 619–629. - PubMed
-
- Azevedo, C. , Sadanandom, A. , Kitagawa, K. , Freialdenhoven, A. , Shirasu, K. and Schulze‐Lefert, P. (2002) The RAR1 interactor SGT1, an essential component of R gene‐triggered disease resistance. Science, 295, 2073–3016. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
