

Opportunities and challenges in the therapeutic activation of human energy expenditure and thermogenesis to manage obesity
- PMID: 31914415
- PMCID: PMC7029124
- DOI: 10.1074/jbc.REV119.007363
Opportunities and challenges in the therapeutic activation of human energy expenditure and thermogenesis to manage obesity
Abstract
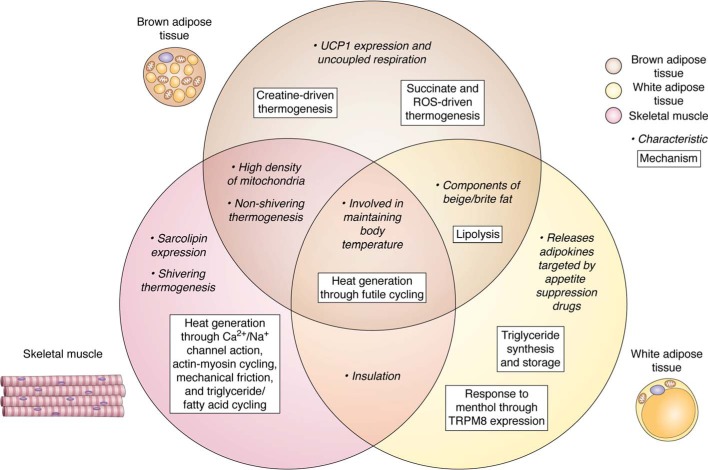
The current obesity pandemic results from a physiological imbalance in which energy intake chronically exceeds energy expenditure (EE), and prevention and treatment strategies remain generally ineffective. Approaches designed to increase EE have been informed by decades of experiments in rodent models designed to stimulate adaptive thermogenesis, a long-term increase in metabolism, primarily induced by chronic cold exposure. At the cellular level, thermogenesis is achieved through increased rates of futile cycling, which are observed in several systems, most notably the regulated uncoupling of oxidative phosphorylation from ATP generation by uncoupling protein 1, a tissue-specific protein present in mitochondria of brown adipose tissue (BAT). Physiological activation of BAT and other organ thermogenesis occurs through β-adrenergic receptors (AR), and considerable effort over the past 5 decades has been directed toward developing AR agonists capable of safely achieving a net negative energy balance while avoiding unwanted cardiovascular side effects. Recent discoveries of other BAT futile cycles based on creatine and succinate have provided additional targets. Complicating the current and developing pharmacological-, cold-, and exercise-based methods to increase EE is the emerging evidence for strong physiological drives toward restoring lost weight over the long term. Future studies will need to address technical challenges such as how to accurately measure individual tissue thermogenesis in humans; how to safely activate BAT and other organ thermogenesis; and how to sustain a negative energy balance over many years of treatment.
Keywords: adipocyte; adrenergic receptor; energy expenditure; energy metabolism; imaging; metabolic disorder; obesity; pharmacology; sympathomimetic; thermogenesis.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures



References
-
- Schauer P. R., Bhatt D. L., Kirwan J. P., Wolski K., Brethauer S. A., Navaneethan S. D., Aminian A., Pothier C. E., Kim E. S., Nissen S. E., Kashyap S. R., and STAMPEDE Investigators (2014) Bariatric surgery versus intensive medical therapy for diabetes–3-year outcomes. N. Engl. J. Med. 370, 2002–2013 10.1056/NEJMoa1401329 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
