

A promiscuous inflammasome sparks replication of a common tumor virus
- PMID: 31919284
- PMCID: PMC6983388
- DOI: 10.1073/pnas.1919133117
A promiscuous inflammasome sparks replication of a common tumor virus
Abstract
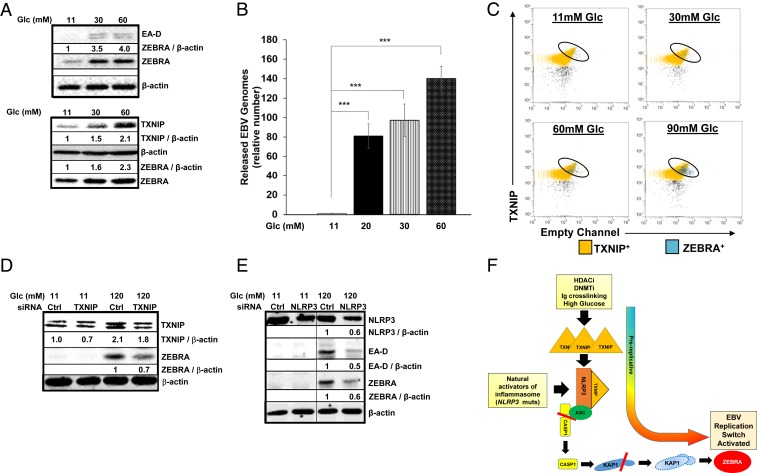
Viruses activate inflammasomes but then subvert resulting inflammatory responses to avoid elimination. We asked whether viruses could instead use such activated or primed inflammasomes to directly aid their propagation and spread. Since herpesviruses are experts at coopting cellular functions, we investigated whether Epstein-Barr virus (EBV), an oncoherpesvirus, exploits inflammasomes to activate its replicative or lytic phase. Indeed, our experiments reveal that EBV exploits several inflammasome sensors to actually activate its replicative phase from quiescence/latency. In particular, TXNIP, a key inflammasome intermediary, causes assembly of the NLRP3 inflammasome, resulting in caspase-1-mediated depletion of the heterochromatin-inducing epigenetic repressor KAP1/TRIM28 in a subpopulation of cells. As a result, only TXNIPhiKAP1lo cells, that is, in a primed/prolytic state, turn expression of the replication/lytic/reactivation switch protein on to enter the replicative phase. Our findings 1) demonstrate that EBV dovetails its escape strategy to a key cellular danger-sensing mechanism, 2) indicate that transcription may be regulated by KAP1 abundance aside from canonical regulation through its posttranslational modification, 3) mechanistically link diabetes, which frequently activates the NLRP3 inflammasome, to deregulation of a tumor virus, and 4) demonstrate that B lymphocytes from NOMID (neonatal onset multisystem inflammatory disease) patients who have NLRP3 mutations and suffer from hyperactive innate responses are defective in controlling a herpesvirus.
Keywords: Epstein−Barr virus; KAP1; TXNIP; diabetes; inflammasome.
Conflict of interest statement
The authors declare no competing interest.
Figures
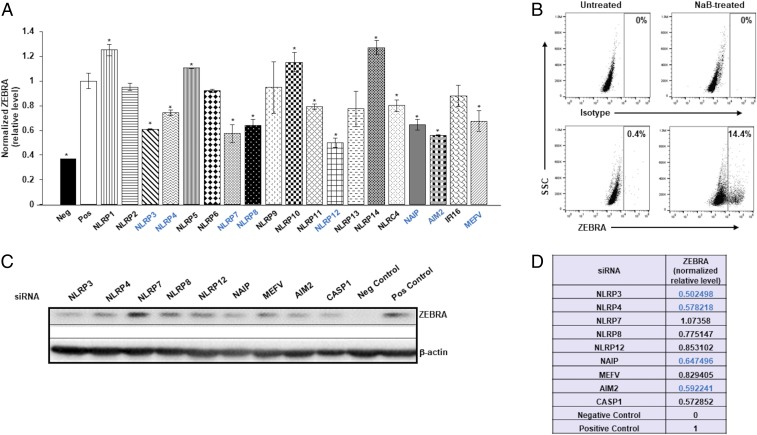
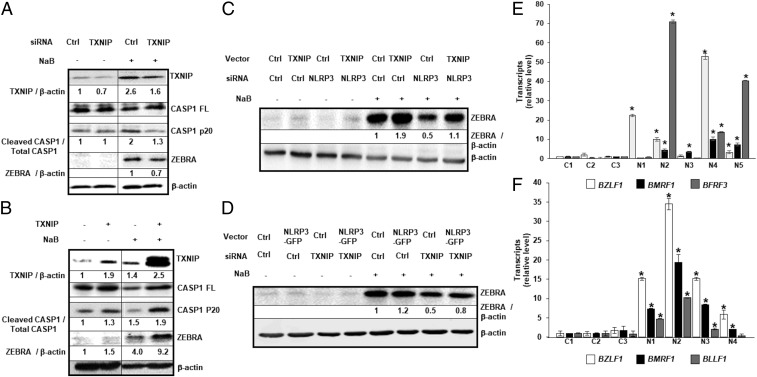
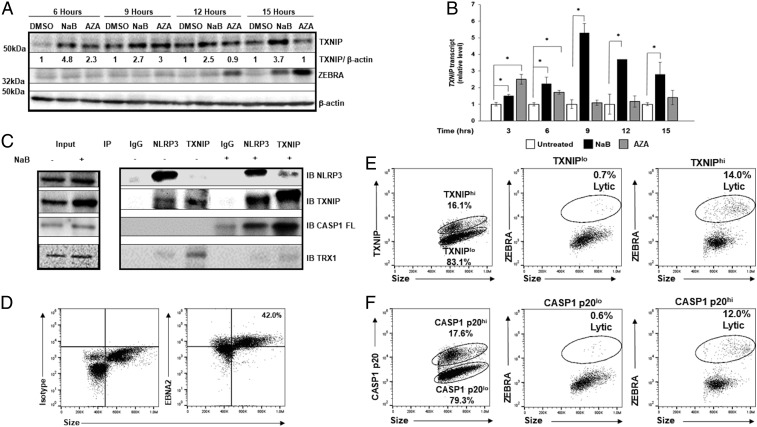
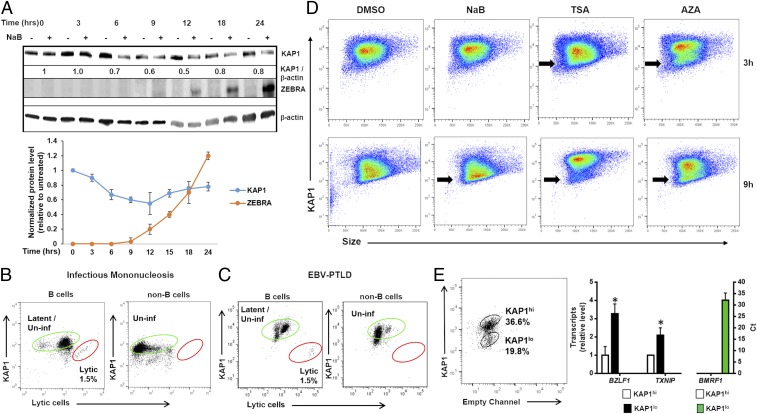
References
-
- Broz P., Dixit V. M., Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 16, 407–420 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
