

Modulation of Transient Receptor Potential C Channel Activity by Cholesterol
- PMID: 31920669
- PMCID: PMC6923273
- DOI: 10.3389/fphar.2019.01487
Modulation of Transient Receptor Potential C Channel Activity by Cholesterol
Abstract
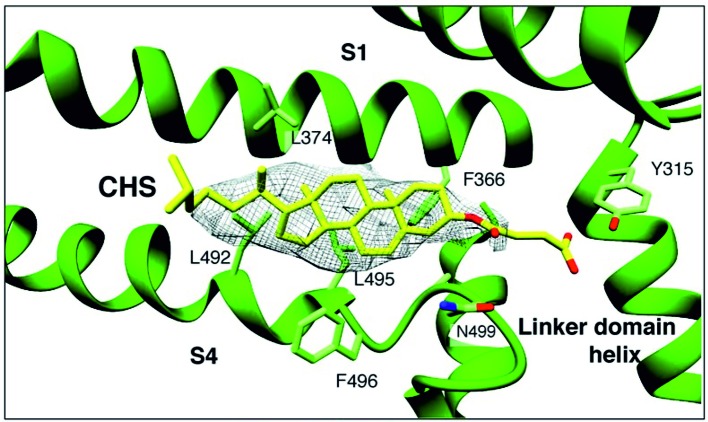
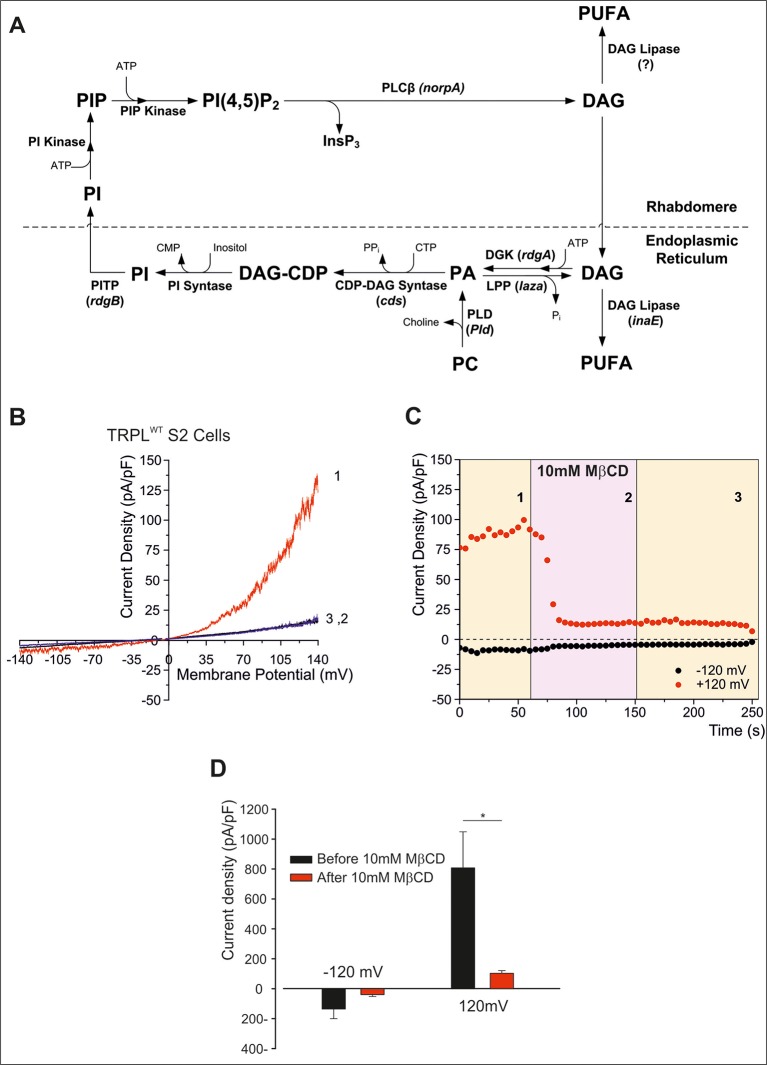
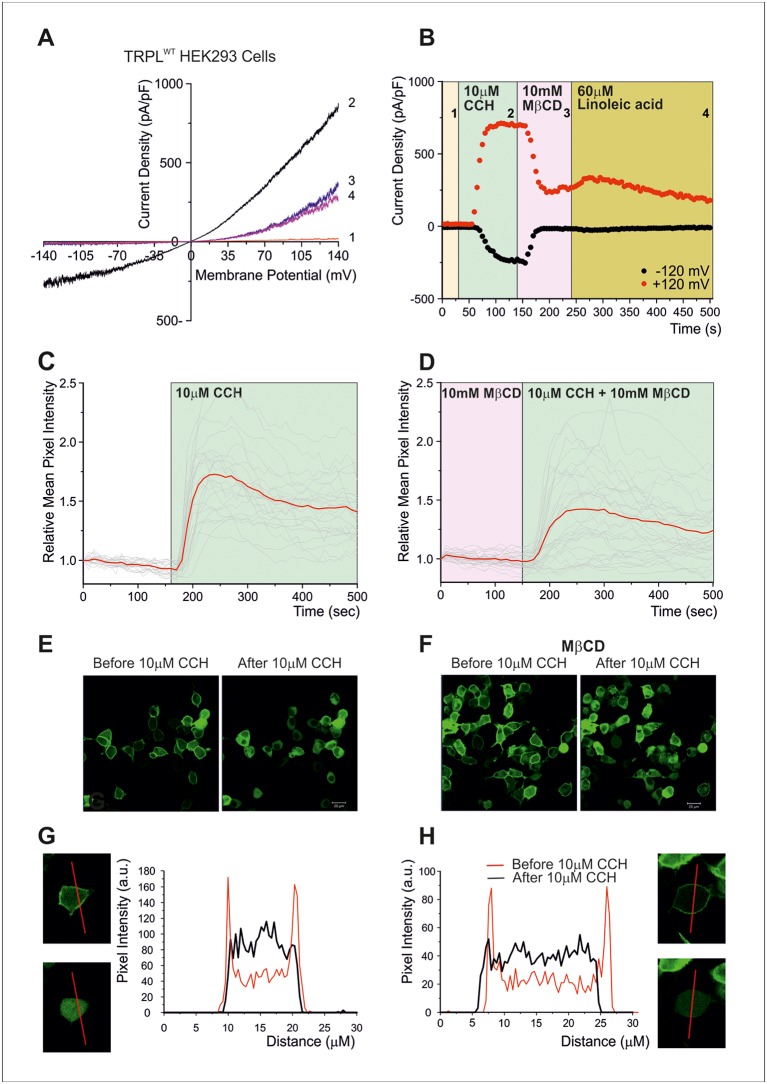
Changes of cholesterol level in the plasma membrane of cells have been shown to modulate ion channel function. The proposed mechanisms underlying these modulations include association of cholesterol to a single binding site at a single channel conformation, association to a highly flexible cholesterol binding site adopting multiple poses, and perturbation of lipid rafts. These perturbations have been shown to induce reversible targeting of mammalian transient receptor potential C (TRPC) channels to the cholesterol-rich membrane environment of lipid rafts. Thus, the observed inhibition of TRPC channels by methyl-β-cyclodextrin (MβCD), which induces cholesterol efflux from the plasma membrane, may result from disruption of lipid rafts. This perturbation was also shown to disrupt multimolecular signaling complexes containing TRPC channels. The Drosophila TRP and TRP-like (TRPL) channels belong to the TRPC channel subfamily. When the Drosophila TRPL channel was expressed in S2 or HEK293 cells and perfused with MβCD, the TRPL current was abolished in less than 100 s, fitting well the fast kinetic phase of cholesterol sequestration experiments in cells. It was thus suggested that the fast kinetics of TRPL channel suppression by MβCD arise from disruption of lipid rafts. Accordingly, lipid raft perturbation by cholesterol sequestration could give clues to the function of lipid environment in TRPC channel activity and its mechanism.
Keywords: TRP-like (TRPL) channel; caveolae; cholesterol recognition amino acid consensus sequence (CRAC); lipid rafts; methyl-β-cyclodextrin.
Copyright © 2019 Gutorov, Peters, Katz, Brandwine, Barbera, Levitan and Minke.
Figures
References
-
- Adebiyi A., Narayanan D., Jaggar J. H. (2011). Caveolin-1 assembles type 1 inositol 1,4,5-trisphosphate receptors and canonical transient receptor potential 3 channels into a functional signaling complex in arterial smooth muscle cells. J. Biol. Chem. 286, 4341–4348. 10.1074/jbc.M110.179747 - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases