

Emerging Mechanisms of Insulin-Mediated Antiviral Immunity in Drosophila melanogaster
- PMID: 31921210
- PMCID: PMC6934001
- DOI: 10.3389/fimmu.2019.02973
Emerging Mechanisms of Insulin-Mediated Antiviral Immunity in Drosophila melanogaster
Abstract
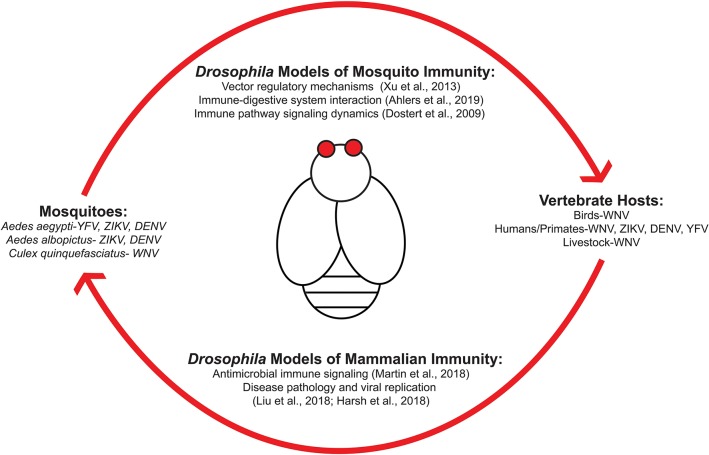
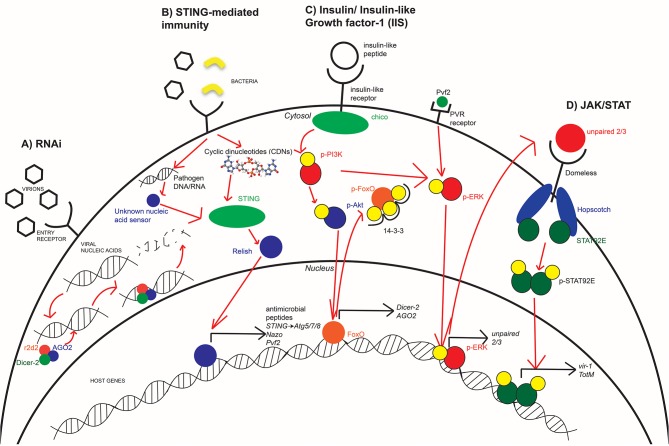
Arboviruses (arthropod-borne viruses), such as Zika (ZIKV), West Nile (WNV), and dengue (DENV) virus, include some of the most significant global health risks to human populations. The steady increase in the number of cases is of great concern due to the debilitating diseases associated with each viral infection. Because these viruses all depend on the mosquito as a vector for disease transmission, current research has focused on identifying immune mechanisms used by insects to effectively harbor these viruses and cause disease in humans and other animals. Drosophila melanogaster are a vital model to study arboviral infections and host responses as they are a genetically malleable model organism for experimentation that can complement analysis in the virus' natural vectors. D. melanogaster encode a number of distinct mechanisms of antiviral defense that are found in both mosquito and vertebrate animal systems, providing a viable model for study. These pathways include canonical antiviral modules such as RNA interference (RNAi), JAK/STAT signaling, and the induction of STING-mediated immune responses like autophagy. Insulin signaling plays a significant role in host-pathogen interactions. The exact mechanisms of insulin-mediated immune responses vary with each virus type, but nevertheless ultimately demonstrates that metabolic and immune signaling are coupled for antiviral immunity in an arthropod model. This mini review provides our current understanding of antiviral mechanisms in D. melanogaster, with a focus on insulin-mediated antiviral signaling, and how such immune responses pertain to disease models in vertebrate and mosquito species.
Keywords: JAK/STAT; RNA interference; STING; West Nile virus; Zika virus; dengue virus; innate immunity; insulin.
Copyright © 2019 Trammell and Goodman.
Figures
References
-
- Muttis E, Balsalobre A, Chuchuy A, Mangudo C, Ciota AT, Kramer LD, et al. . Factors related to Aedes aegypti (Diptera: Culicidae) populations and temperature determine differences on life-history traits with regional implications in disease transmission. J Med Entomol. (2018) 55:1105–12. 10.1093/jme/tjy057 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials