

Inflorescence Meristem Fate Is Dependent on Seed Development and FRUITFULL in Arabidopsis thaliana
- PMID: 31921264
- PMCID: PMC6930240
- DOI: 10.3389/fpls.2019.01622
Inflorescence Meristem Fate Is Dependent on Seed Development and FRUITFULL in Arabidopsis thaliana
Abstract
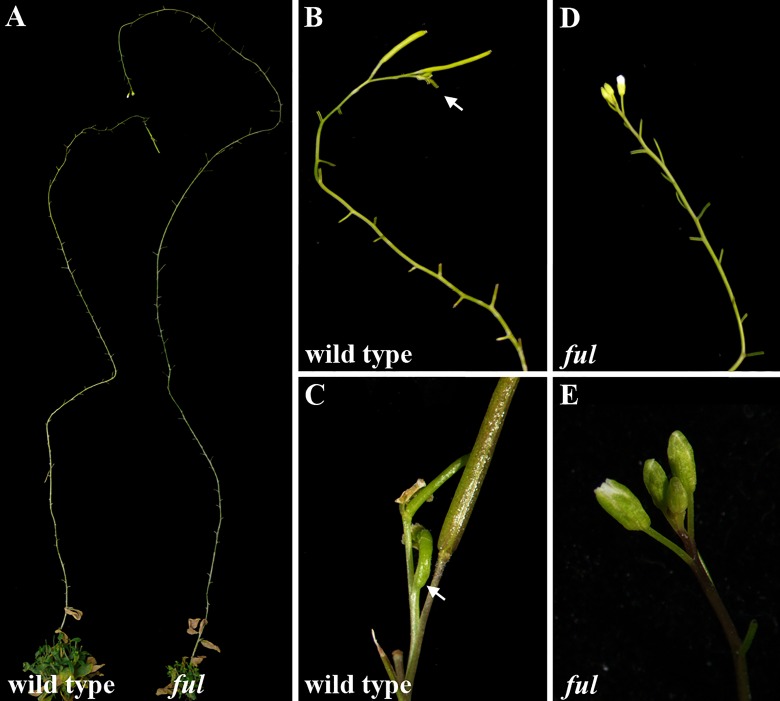
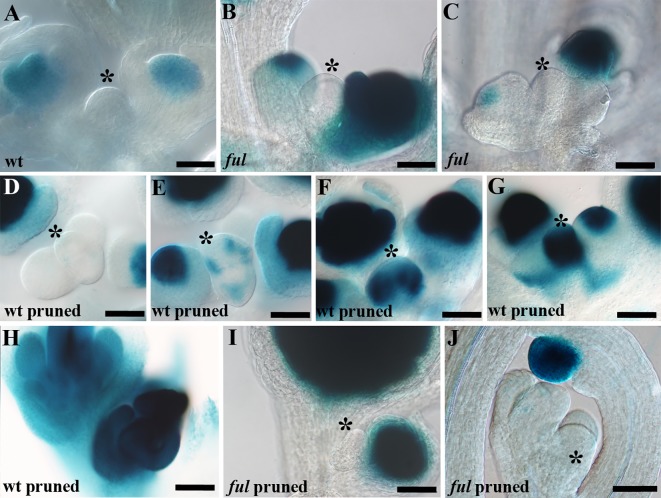
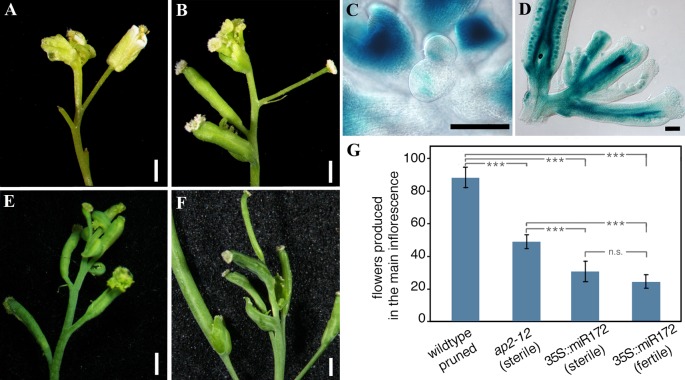
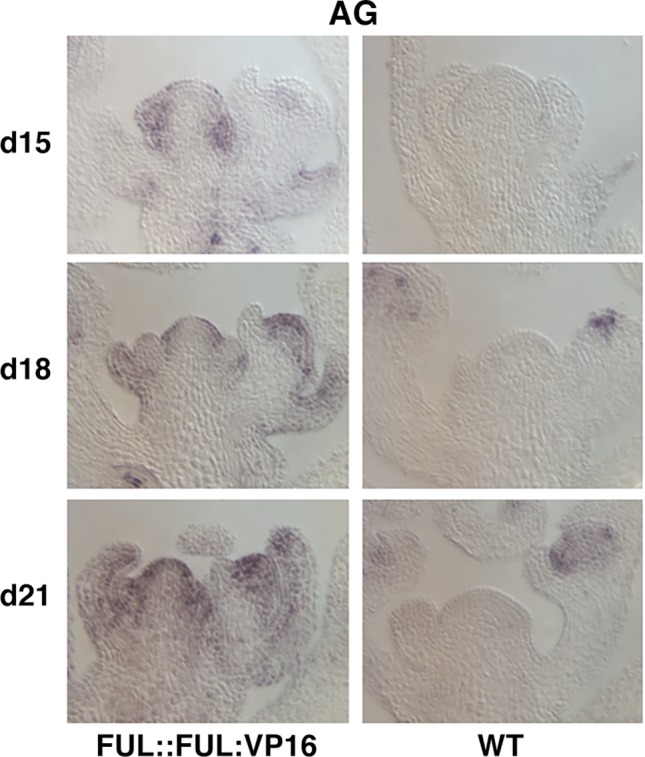
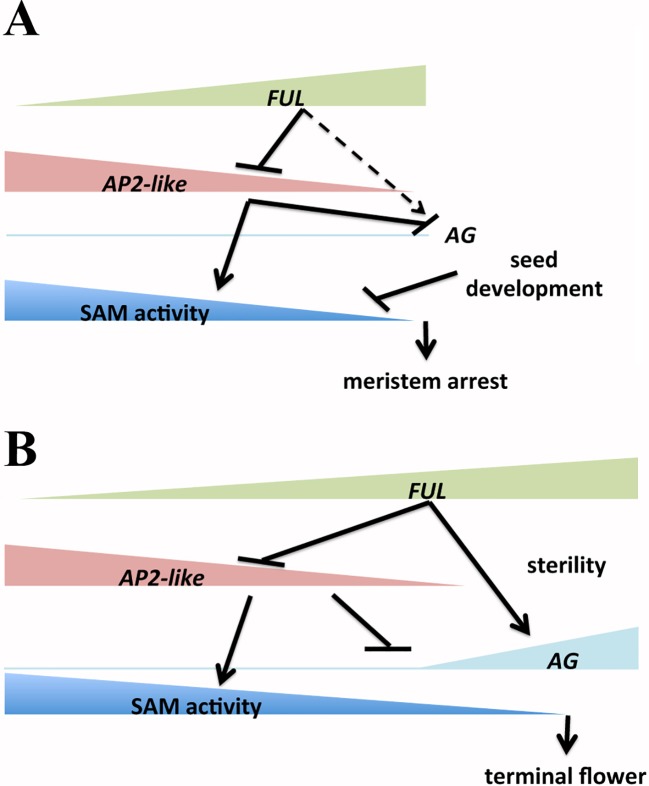
After a vegetative phase, plants initiate the floral transition in response to both environmental and endogenous cues to optimize reproductive success. During this process, the vegetative shoot apical meristem (SAM), which was producing leaves and branches, becomes an inflorescence SAM and starts producing flowers. Inflorescences can be classified in two main categories, depending on the fate of the inflorescence meristem: determinate or indeterminate. In determinate inflorescences, the SAM differentiates directly, or after the production of a certain number of flowers, into a flower, while in indeterminate inflorescences the SAM remains indeterminate and produces continuously new flowers. Even though indeterminate inflorescences have an undifferentiated SAM, the number of flowers produced by a plant is not indefinite and is characteristic of each species, indicating that it is under genetic control. In Arabidopsis thaliana and other species with indeterminate inflorescences, the end of flower production occurs by a regulated proliferative arrest of inflorescence meristems on all reproductive branches that is reminiscent of a state of induced dormancy and does not involve the determination of the SAM. This process is controlled genetically by the FRUITFULL-APETALA2 (FUL-AP2) pathway and by a correlative control exerted by the seeds through a mechanism not well understood yet. In the absence of seeds, meristem proliferative arrest does not occur, and the SAM remains actively producing flowers until it becomes determinate, differentiating into a terminal floral structure. Here we show that the indeterminate growth habit of Arabidopsis inflorescences is a facultative condition imposed by the meristematic arrest directed by FUL and the correlative signal of seeds. The terminal differentiation of the SAM when seed production is absent correlates with the induction of AGAMOUS expression in the SAM. Moreover, terminal flower formation is strictly dependent on the activity of FUL, as it was never observed in ful mutants, regardless of the fertility of the plant or the presence/absence of the AG repression exerted by APETALA2 related factors.
Keywords: AGAMOUS; FRUITFULL; inflorescence development; inflorescence meristem; inflorescence proliferative arrest; terminal flower.
Copyright © 2019 Balanzà, Martínez-Fernández, Sato, Yanofsky and Ferrándiz.
Figures


References
-
- Alvarez J., Smyth D. R. (1999). CRABS CLAW and SPATULA, two Arabidopsis genes that control carpel development in parallel with AGAMOUS. Development 126 (11), 2377–2386. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials