

Redox Balance Keepers and Possible Cell Functions Managed by Redox Homeostasis in Trypanosoma cruzi
- PMID: 31921709
- PMCID: PMC6932984
- DOI: 10.3389/fcimb.2019.00435
Redox Balance Keepers and Possible Cell Functions Managed by Redox Homeostasis in Trypanosoma cruzi
Abstract
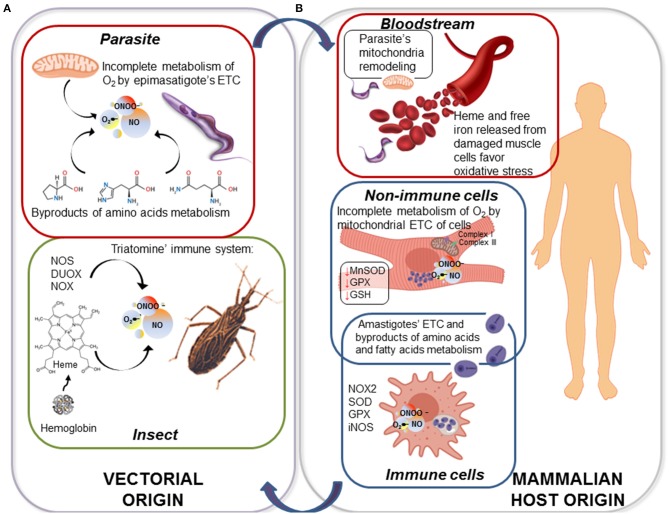
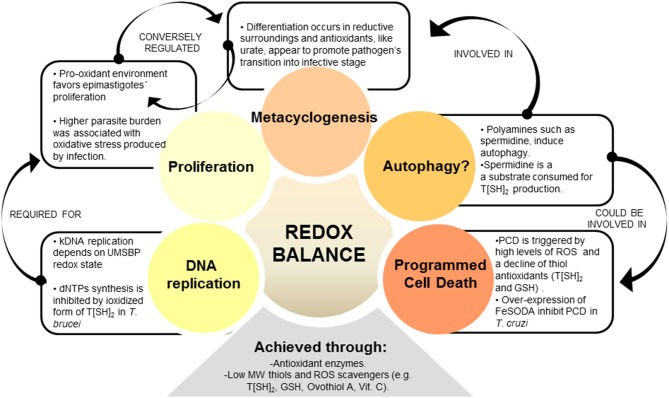
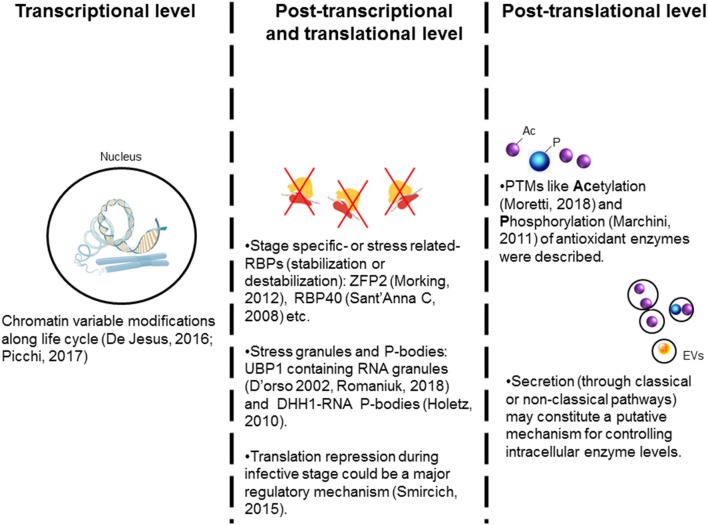
The toxicity of oxygen and nitrogen reactive species appears to be merely the tip of the iceberg in the world of redox homeostasis. Now, oxidative stress can be seen as a two-sided process; at high concentrations, it causes damage to biomolecules, and thus, trypanosomes have evolved a strong antioxidant defense system to cope with these stressors. At low concentrations, oxidants are essential for cell signaling, and in fact, the oxidants/antioxidants balance may be able to trigger different cell fates. In this comprehensive review, we discuss the current knowledge of the oxidant environment experienced by T. cruzi along the different phases of its life cycle, and the molecular tools exploited by this pathogen to deal with oxidative stress, for better or worse. Further, we discuss the possible redox-regulated processes that could be governed by this oxidative context. Most of the current research has addressed the importance of the trypanosomes' antioxidant network based on its detox activity of harmful species; however, new efforts are necessary to highlight other functions of this network and the mechanisms underlying the fine regulation of the defense machinery, as this represents a master key to hinder crucial pathogen functions. Understanding the relevance of this balance keeper program in parasite biology will give us new perspectives to delineate improved treatment strategies.
Keywords: Trypanosoma cruzi; antioxidant network; redox-dependent mechanisms; regulation; stage-specific oxidants.
Copyright © 2019 Mesías, Garg and Zago.
Figures
References
-
- Alvarez M. N., Peluffo G., Piacenza L., Radi R. (2011). Intraphagosomal peroxynitrite as a macrophage-derived cytotoxin against internalized Trypanosoma cruzi: consequences for oxidative killing and role of microbial peroxiredoxins in infectivity. J. Biol. Chem. 286, 6627–6640. 10.1074/jbc.M110.167247 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical