

Astrocyte Unfolded Protein Response Induces a Specific Reactivity State that Causes Non-Cell-Autonomous Neuronal Degeneration
- PMID: 31924446
- PMCID: PMC7054837
- DOI: 10.1016/j.neuron.2019.12.014
Astrocyte Unfolded Protein Response Induces a Specific Reactivity State that Causes Non-Cell-Autonomous Neuronal Degeneration
Abstract
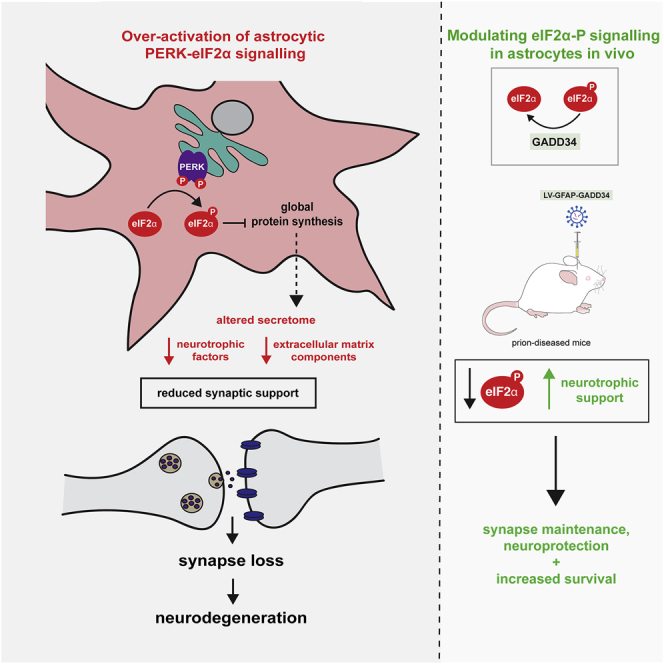
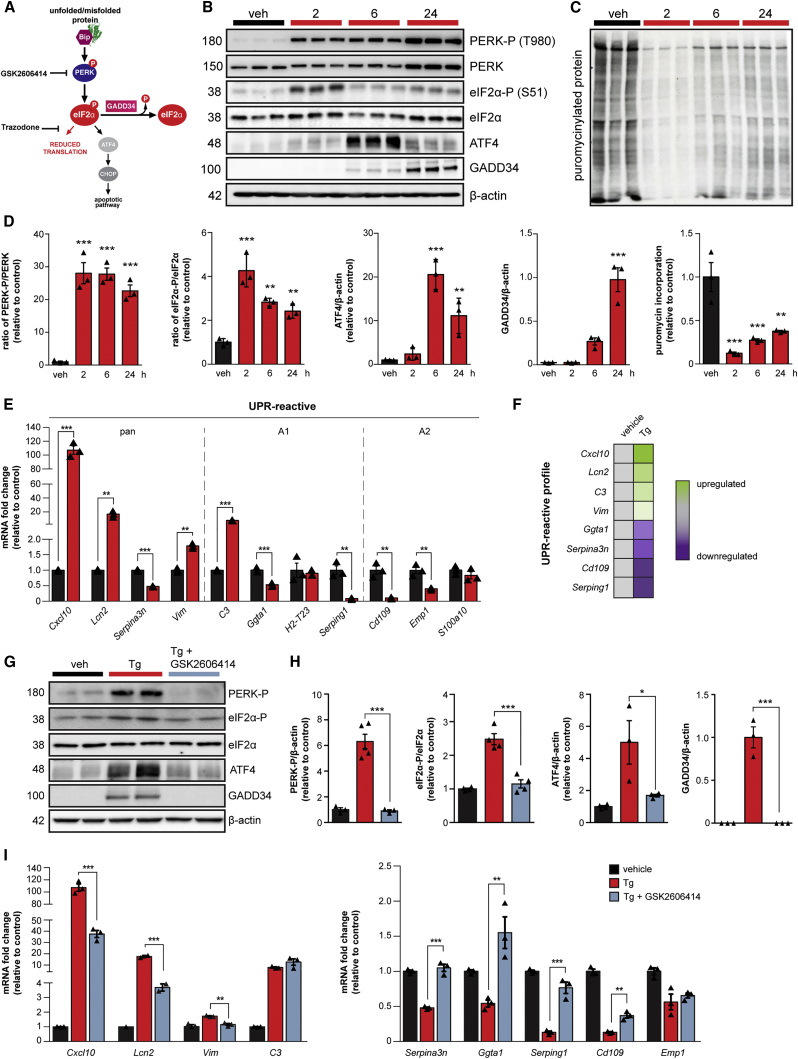
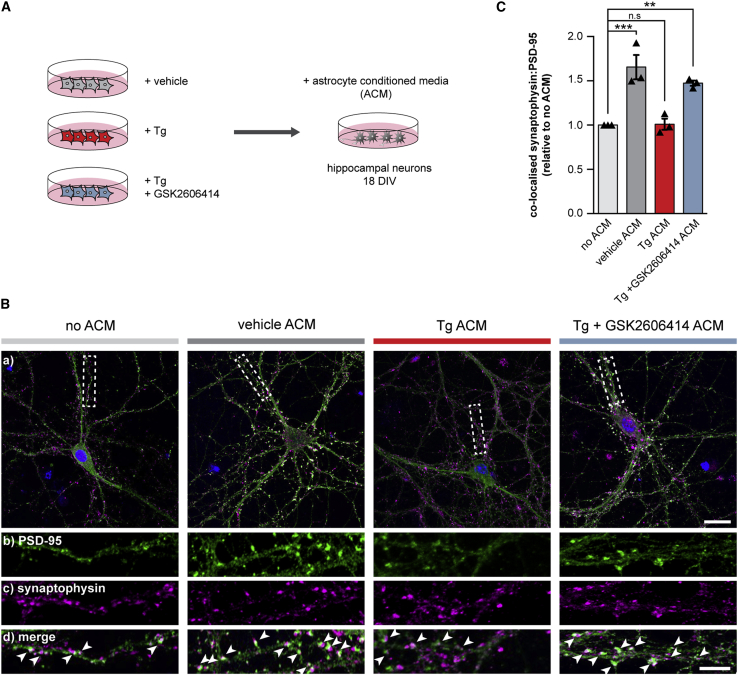
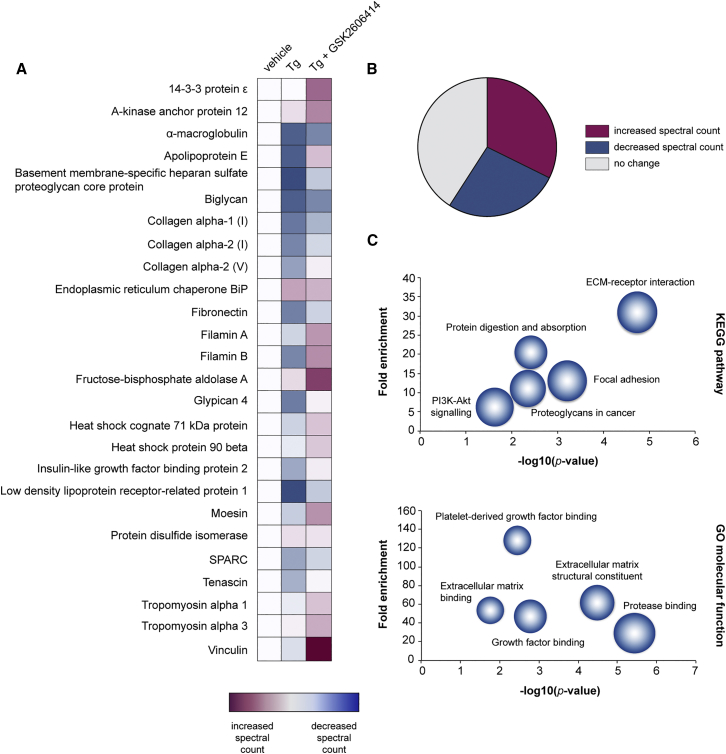
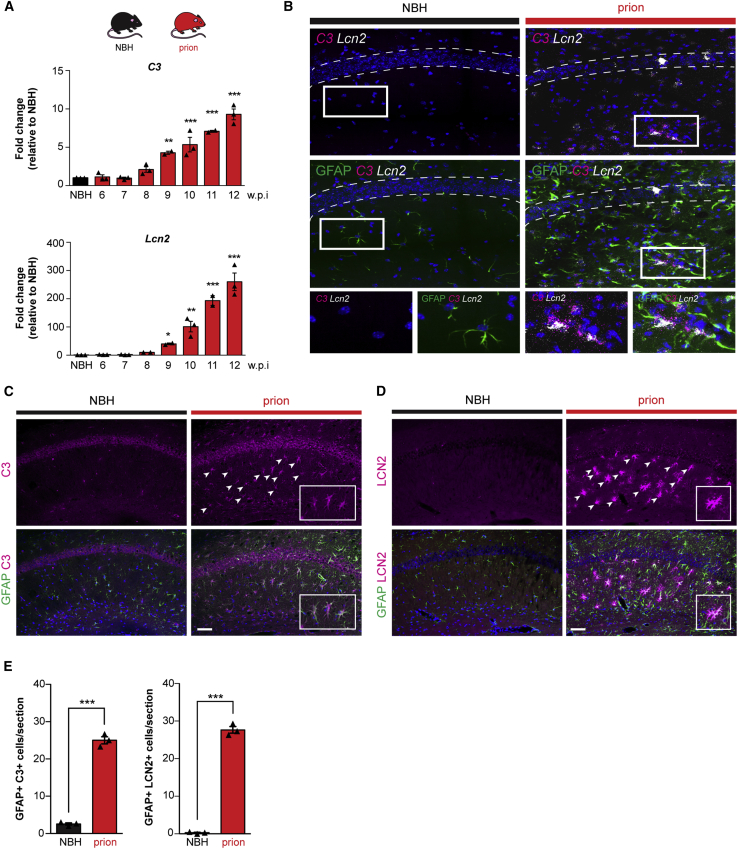
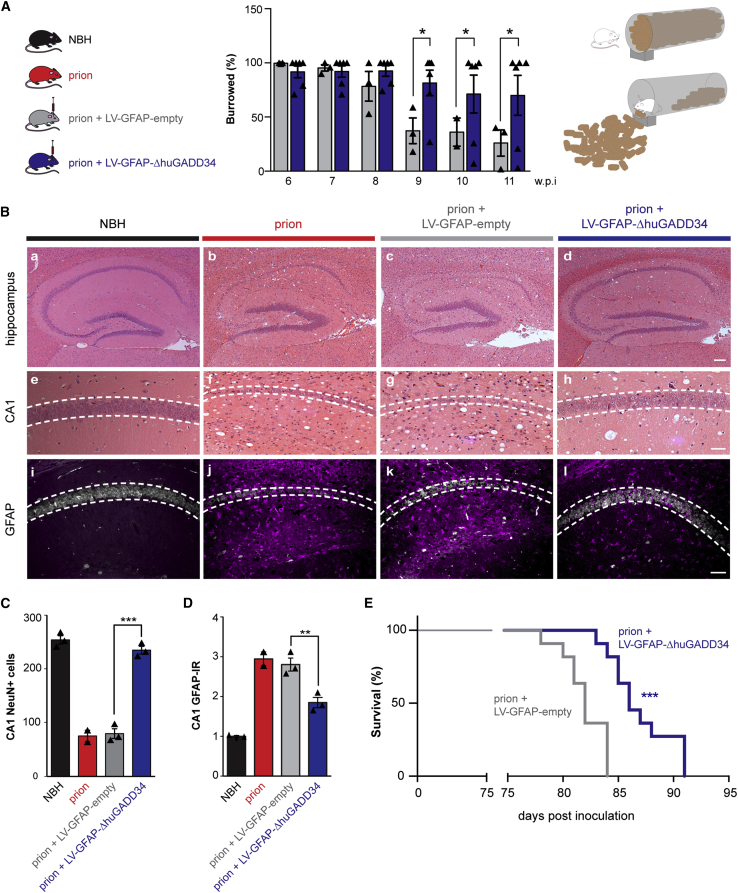
Recent interest in astrocyte activation states has raised the fundamental question of how these cells, normally essential for synapse and neuronal maintenance, become pathogenic. Here, we show that activation of the unfolded protein response (UPR), specifically phosphorylated protein kinase R-like endoplasmic reticulum (ER) kinase (PERK-P) signaling-a pathway that is widely dysregulated in neurodegenerative diseases-generates a distinct reactivity state in astrocytes that alters the astrocytic secretome, leading to loss of synaptogenic function in vitro. Further, we establish that the same PERK-P-dependent astrocyte reactivity state is harmful to neurons in vivo in mice with prion neurodegeneration. Critically, targeting this signaling exclusively in astrocytes during prion disease is alone sufficient to prevent neuronal loss and significantly prolongs survival. Thus, the astrocyte reactivity state resulting from UPR over-activation is a distinct pathogenic mechanism that can by itself be effectively targeted for neuroprotection.
Keywords: LCN2; PERK signalling; astrocyte reactivity state; astrocytes; neurodegeneration; neuroprotection; secretome; synapse; translational neuroscience; unfolded protein response.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests. O.J.F. is now an employee of AstraZeneca.
Figures

Comment in
-
Withdrawn support.Nat Rev Neurosci. 2020 Mar;21(3):119. doi: 10.1038/s41583-020-0268-4. Nat Rev Neurosci. 2020. PMID: 31996802 No abstract available.
References
-
- Axten J.M., Medina J.R., Feng Y., Shu A., Romeril S.P., Grant S.W., Li W.H., Heerding D.A., Minthorn E., Mencken T. Discovery of 7-methyl-5-(1-{[3-(trifluoromethyl)phenyl]acetyl}-2,3-dihydro-1H-indol-5-yl)-7H-pyrrolo[2,3-d]pyrimidin-4-amine (GSK2606414), a potent and selective first-in-class inhibitor of protein kinase R (PKR)-like endoplasmic reticulum kinase (PERK) J. Med. Chem. 2012;55:7193–7207. - PubMed
-
- Christopherson K.S., Ullian E.M., Stokes C.C., Mullowney C.E., Hell J.W., Agah A., Lawler J., Mosher D.F., Bornstein P., Barres B.A. Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell. 2005;120:421–433. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MC_PC_17230/MRC_/Medical Research Council/United Kingdom
- G0300336/MRC_/Medical Research Council/United Kingdom
- G0701476/MRC_/Medical Research Council/United Kingdom
- MC_U132692719/MRC_/Medical Research Council/United Kingdom
- MR/K008803/1/MRC_/Medical Research Council/United Kingdom
- G0802545/MRC_/Medical Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- MR/M010503/1/MRC_/Medical Research Council/United Kingdom
- MC_PC_12009/MRC_/Medical Research Council/United Kingdom
- 201487/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- MR/R024820/1/MRC_/Medical Research Council/United Kingdom
- MR/S00503X/1/MRC_/Medical Research Council/United Kingdom
- G0300338/MRC_/Medical Research Council/United Kingdom
- G0700392/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
