

Single-molecule imaging analysis reveals the mechanism of a high-catalytic-activity mutant of chitinase A from Serratia marcescens
- PMID: 31924658
- PMCID: PMC7029130
- DOI: 10.1074/jbc.RA119.012078
Single-molecule imaging analysis reveals the mechanism of a high-catalytic-activity mutant of chitinase A from Serratia marcescens
Abstract
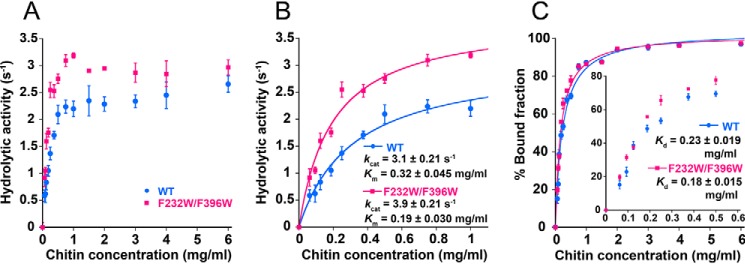
Chitin degradation is important for biomass conversion and has potential applications for agriculture, biotechnology, and the pharmaceutical industry. Chitinase A from the Gram-negative bacterium Serratia marcescens (SmChiA) is a processive enzyme that hydrolyzes crystalline chitin as it moves linearly along the substrate surface. In a previous study, the catalytic activity of SmChiA against crystalline chitin was found to increase after the tryptophan substitution of two phenylalanine residues (F232W and F396W), located at the entrance and exit of the substrate binding cleft of the catalytic domain, respectively. However, the mechanism underlying this high catalytic activity remains elusive. In this study, single-molecule fluorescence imaging and high-speed atomic force microscopy were applied to understand the mechanism of this high-catalytic-activity mutant. A reaction scheme including processive catalysis was used to reproduce the properties of SmChiA WT and F232W/F396W, in which all of the kinetic parameters were experimentally determined. High activity of F232W/F396W mutant was caused by a high processivity and a low dissociation rate constant after productive binding. The turnover numbers for both WT and F232W/F396W, determined by the biochemical analysis, were well-replicated using the kinetic parameters obtained from single-molecule imaging analysis, indicating the validity of the reaction scheme. Furthermore, alignment of amino acid sequences of 258 SmChiA-like proteins revealed that tryptophan, not phenylalanine, is the predominant amino acid at the corresponding positions (Phe-232 and Phe-396 for SmChiA). Our study will be helpful for understanding the kinetic mechanisms and further improvement of crystalline chitin hydrolytic activity of SmChiA mutants.
Keywords: Serratia marcescens; biomass conversion; biotechnology; chitin; chitin degradation; chitinase; high-speed atomic force microscopy; processivity; single-molecule biophysics; single-molecule fluorescence imaging.
© 2020 Visootsat et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures





References
-
- Hamed I., Özogul F., and Regenstein J. M. (2016) Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): a review. Trends Food Sci. Technol. 48, 40–50 10.1016/j.tifs.2015.11.007 - DOI
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
