

Inhibition within a premotor circuit controls the timing of vocal turn-taking in zebra finches
- PMID: 31924758
- PMCID: PMC6954284
- DOI: 10.1038/s41467-019-13938-0
Inhibition within a premotor circuit controls the timing of vocal turn-taking in zebra finches
Abstract
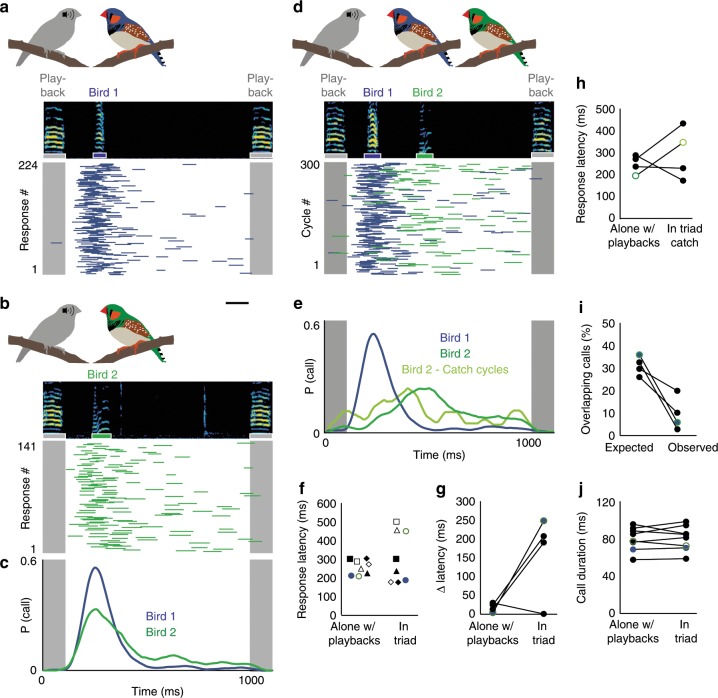
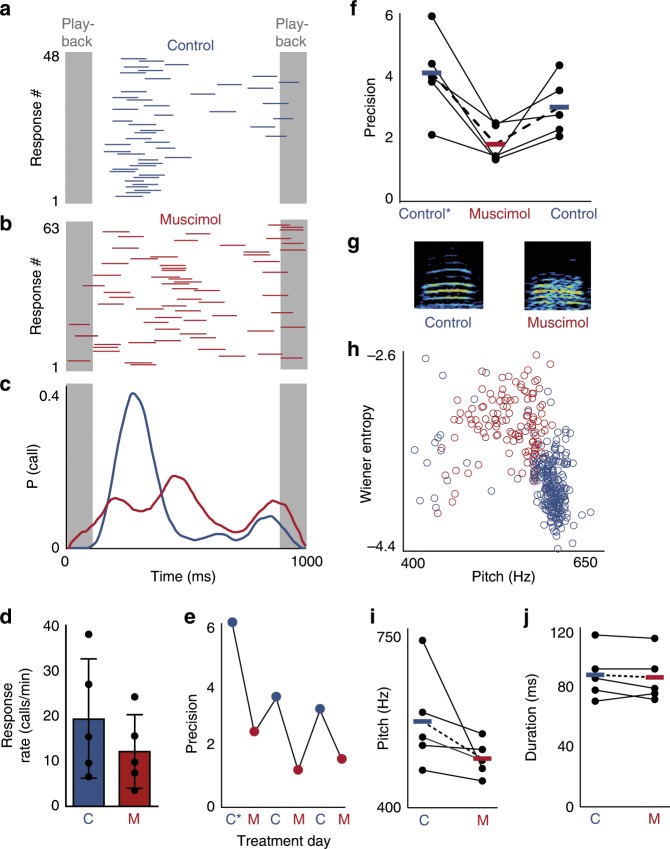
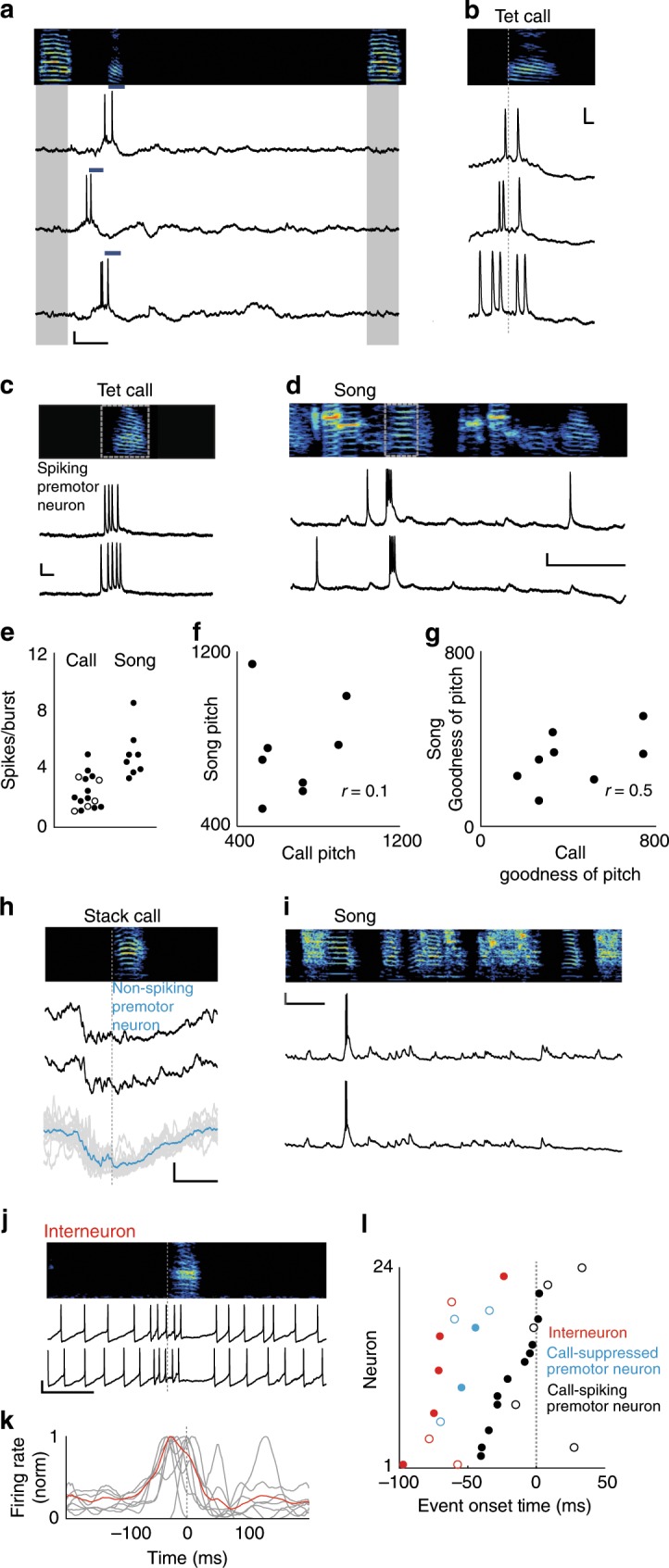
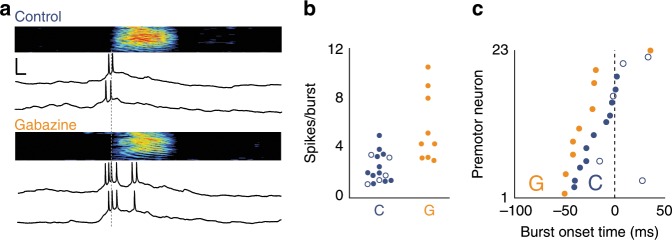
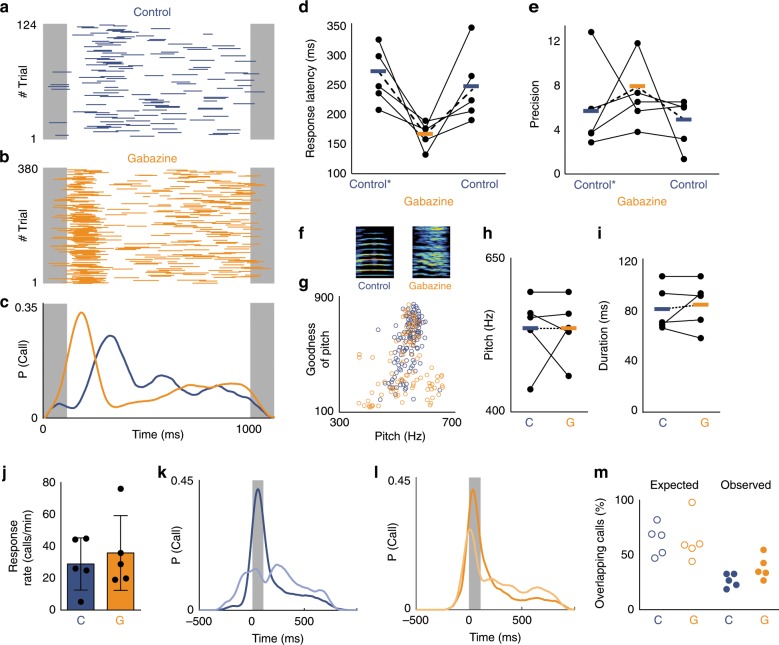
Vocal turn-taking is a fundamental organizing principle of human conversation but the neural circuit mechanisms that structure coordinated vocal interactions are unknown. The ability to exchange vocalizations in an alternating fashion is also exhibited by other species, including zebra finches. With a combination of behavioral testing, electrophysiological recordings, and pharmacological manipulations we demonstrate that activity within a cortical premotor nucleus orchestrates the timing of calls in socially interacting zebra finches. Within this circuit, local inhibition precedes premotor neuron activation associated with calling. Blocking inhibition results in faster vocal responses as well as an impaired ability to flexibly avoid overlapping with a partner. These results support a working model in which premotor inhibition regulates context-dependent timing of vocalizations and enables the precise interleaving of vocal signals during turn-taking.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Heldner M, Edlund J. Pauses, gaps and overlaps in conversations. J. Phonetics. 2010;38:555–568. doi: 10.1016/j.wocn.2010.08.002. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
