

On the existence and functionality of topologically associating domains
- PMID: 31925403
- PMCID: PMC7567612
- DOI: 10.1038/s41588-019-0561-1
On the existence and functionality of topologically associating domains
Abstract
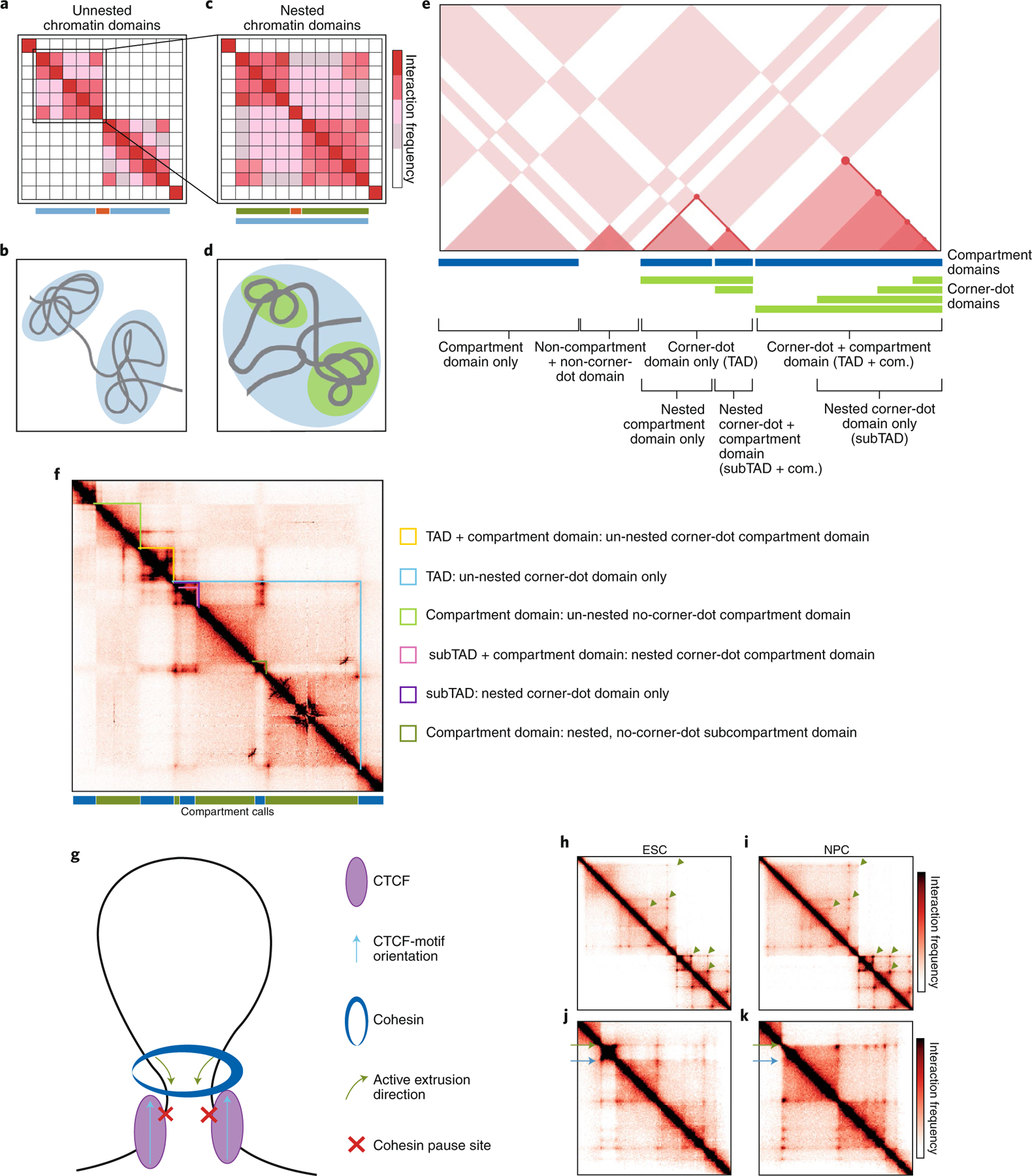
Genomes across a wide range of eukaryotic organisms fold into higher-order chromatin domains. Topologically associating domains (TADs) were originally discovered empirically in low-resolution Hi-C heat maps representing ensemble average interaction frequencies from millions of cells. Here, we discuss recent advances in high-resolution Hi-C, single-cell imaging experiments, and functional genetic studies, which provide an increasingly complex view of the genome's hierarchical structure-function relationship. On the basis of these new findings, we update the definitions of distinct classes of chromatin domains according to emerging knowledge of their structural, mechanistic and functional properties.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures


References
-
- Sexton T et al. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell 148, 458–472 (2012). - PubMed
Publication types
MeSH terms
Grants and funding
- 1DP2MH11024701/U.S. Department of Health & Human Services | NIH | National Institute of Mental Health (NIMH)/International
- R01 MH120269/MH/NIMH NIH HHS/United States
- 1R011MH120269/U.S. Department of Health & Human Services | NIH | National Institute of Mental Health (NIMH)/International
- DP2 MH110247/MH/NIMH NIH HHS/United States
- R37 MH120269/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
