

Investigating eukaryotic cells with cryo-ET
- PMID: 31935172
- PMCID: PMC6960407
- DOI: 10.1091/mbc.E18-05-0329
Investigating eukaryotic cells with cryo-ET
Abstract
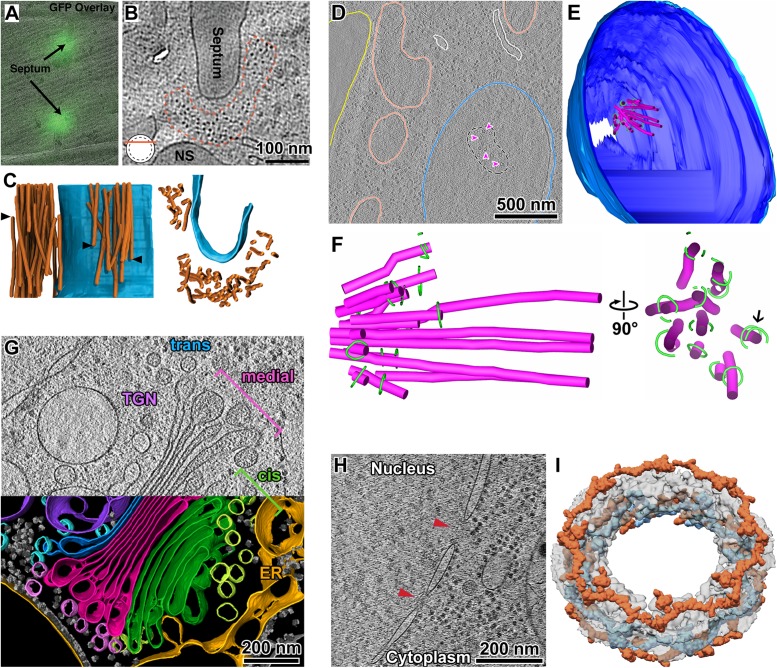
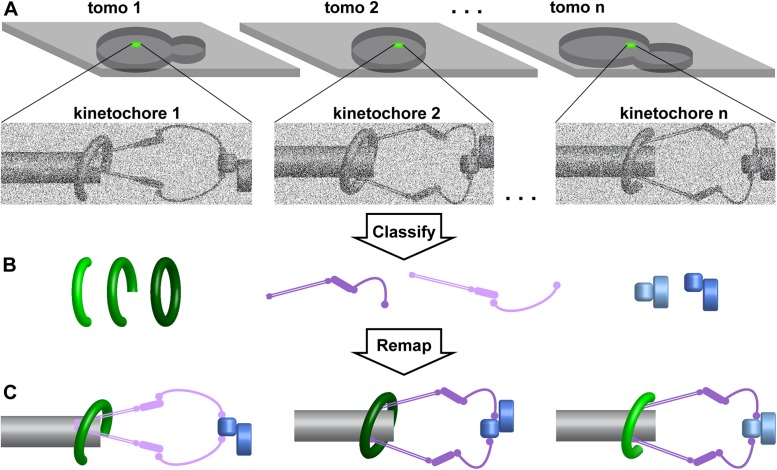
The interior of eukaryotic cells is mysterious. How do the large communities of macromolecular machines interact with each other? How do the structures and positions of these nanoscopic entities respond to new stimuli? Questions like these can now be answered with the help of a method called electron cryotomography (cryo-ET). Cryo-ET will ultimately reveal the inner workings of a cell at the protein, secondary structure, and perhaps even side-chain levels. Combined with genetic or pharmacological perturbation, cryo-ET will allow us to answer previously unimaginable questions, such as how structure, biochemistry, and forces are related in situ. Because it bridges structural biology and cell biology, cryo-ET is indispensable for structural cell biology-the study of the 3-D macromolecular structure of cells. Here we discuss some of the key ideas, strategies, auxiliary techniques, and innovations that an aspiring structural cell biologist will consider when planning to ask bold questions.
Figures




Similar articles
-
Structural investigation of eukaryotic cells: From the periphery to the interior by cryo-electron tomography.Adv Biol Regul. 2023 Jan;87:100923. doi: 10.1016/j.jbior.2022.100923. Epub 2022 Oct 17. Adv Biol Regul. 2023. PMID: 36280452 Review.
-
Cryo-electron tomography for the structural study of mitochondrial translation.Tissue Cell. 2019 Apr;57:129-138. doi: 10.1016/j.tice.2018.08.009. Epub 2018 Aug 29. Tissue Cell. 2019. PMID: 30197222 Review.
-
Developments in cryo-electron tomography for in situ structural analysis.Arch Biochem Biophys. 2015 Sep 1;581:78-85. doi: 10.1016/j.abb.2015.04.006. Epub 2015 Apr 25. Arch Biochem Biophys. 2015. PMID: 25921875 Review.
-
Cryo-electron tomography: gaining insight into cellular processes by structural approaches.Curr Opin Struct Biol. 2011 Oct;21(5):670-7. doi: 10.1016/j.sbi.2011.07.004. Epub 2011 Aug 2. Curr Opin Struct Biol. 2011. PMID: 21813274 Review.
-
In Situ Imaging and Structure Determination of Biomolecular Complexes Using Electron Cryo-Tomography.Methods Mol Biol. 2021;2215:83-111. doi: 10.1007/978-1-0716-0966-8_4. Methods Mol Biol. 2021. PMID: 33368000
Cited by
-
Cryo-EM of a heterogeneous biochemical fraction elucidates multiple protein complexes from a multicellular thermophilic eukaryote.J Struct Biol X. 2023 Aug 9;8:100094. doi: 10.1016/j.yjsbx.2023.100094. eCollection 2023 Dec. J Struct Biol X. 2023. PMID: 37638207 Free PMC article.
-
Heterogeneous non-canonical nucleosomes predominate in yeast cells in situ.Elife. 2023 Jul 28;12:RP87672. doi: 10.7554/eLife.87672. Elife. 2023. PMID: 37503920 Free PMC article.
-
Organelle Interaction and Drug Discovery: Towards Correlative Nanoscopy and Molecular Dynamics Simulation.Front Pharmacol. 2022 Jun 20;13:935898. doi: 10.3389/fphar.2022.935898. eCollection 2022. Front Pharmacol. 2022. PMID: 35795548 Free PMC article. Review.
-
Serial Cryomicrotomy of Saccharomyces cerevisiae for Serial Electron Cryotomography.Bio Protoc. 2020 Nov 20;10(22):e3831. doi: 10.21769/BioProtoc.3831. eCollection 2020 Nov 20. Bio Protoc. 2020. PMID: 33659481 Free PMC article.
-
Cryo-ET detects bundled triple helices but not ladders in meiotic budding yeast.PLoS One. 2022 Apr 14;17(4):e0266035. doi: 10.1371/journal.pone.0266035. eCollection 2022. PLoS One. 2022. PMID: 35421110 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
