

Chronic exposure to methylmercury induces puncta formation in cephalic dopaminergic neurons in Caenorhabditis elegans
- PMID: 31935438
- PMCID: PMC7061079
- DOI: 10.1016/j.neuro.2020.01.003
Chronic exposure to methylmercury induces puncta formation in cephalic dopaminergic neurons in Caenorhabditis elegans
Abstract
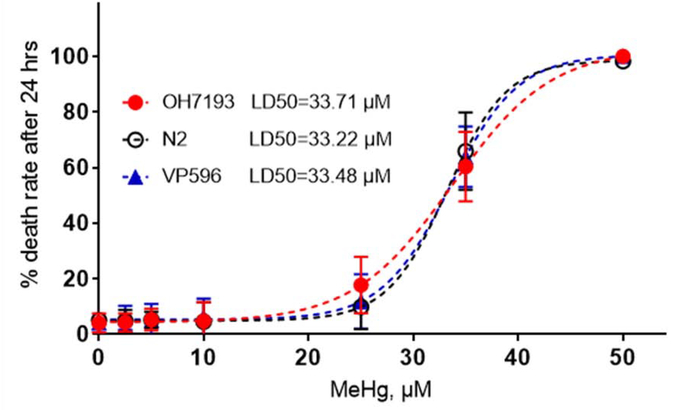
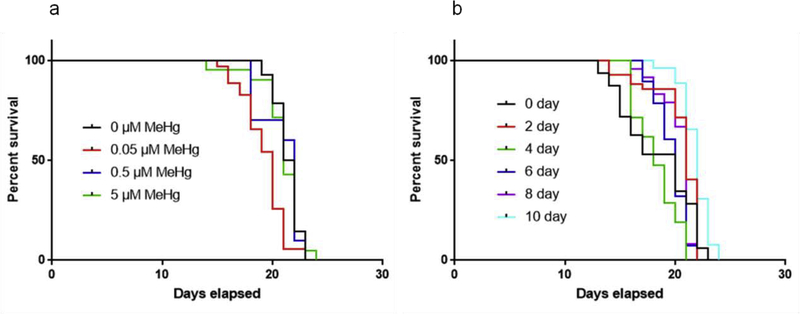
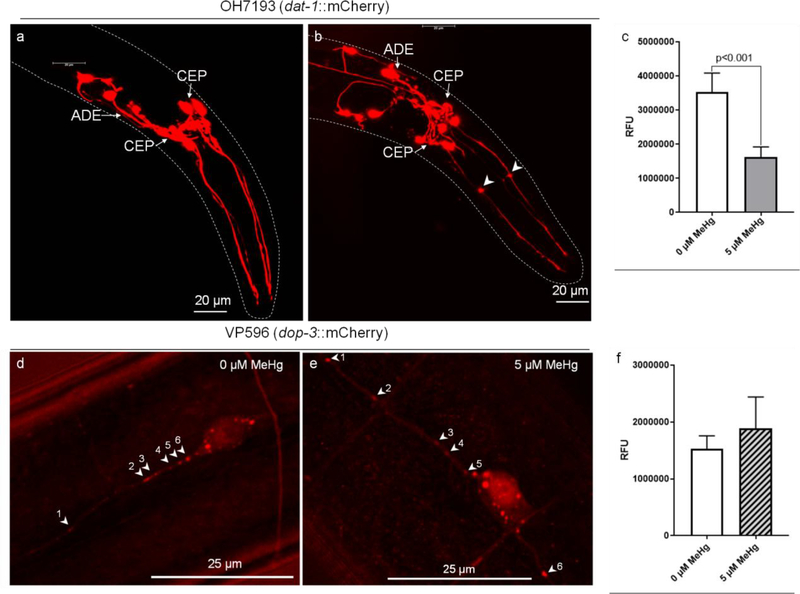
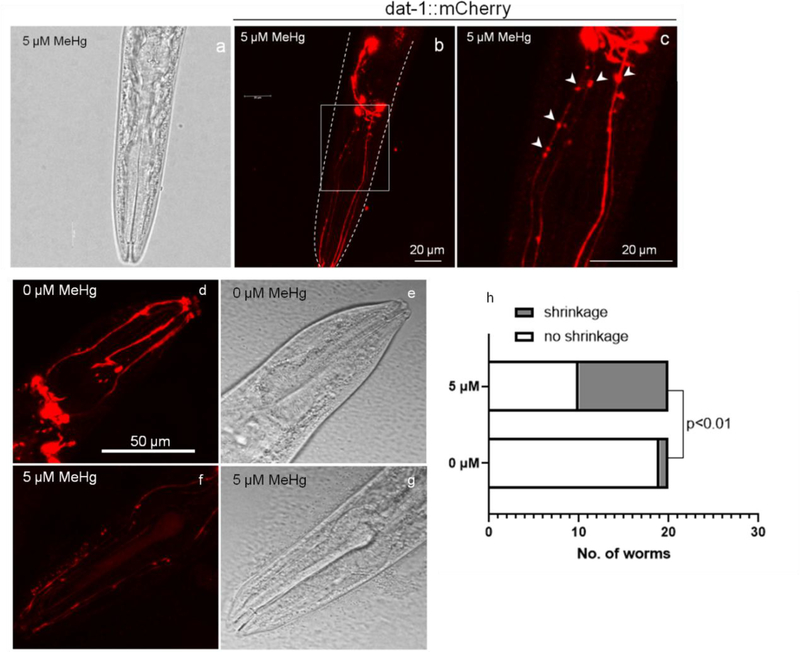
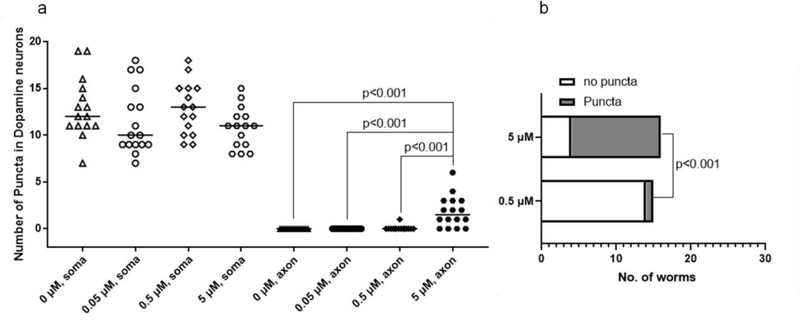
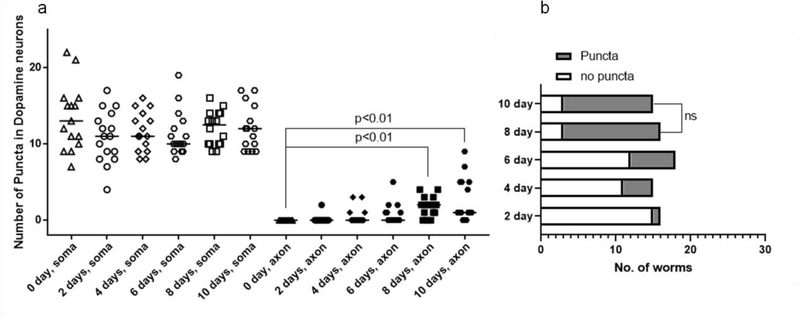
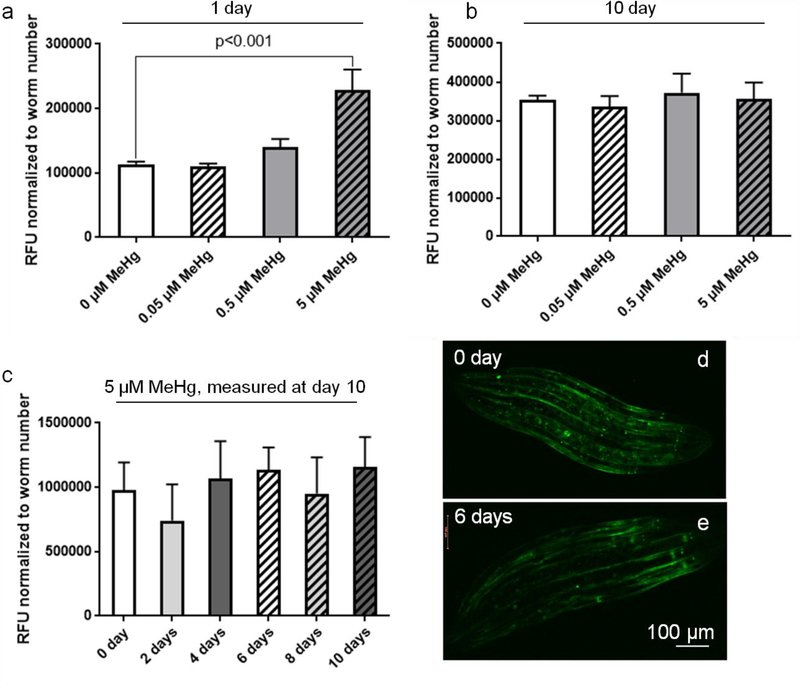
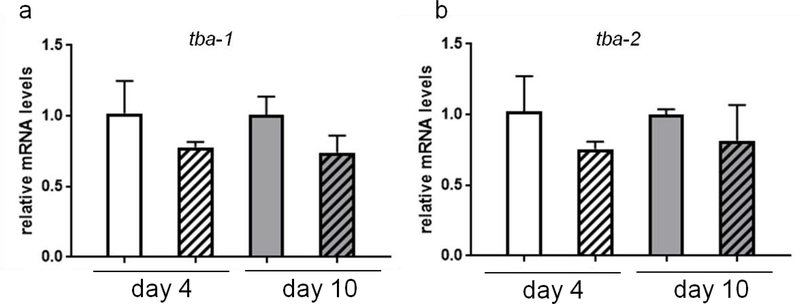
The neurotransmitter dopamine is a neuromodulator in the positive and negative regulation of brain circuits. Dopamine insufficiency or overload has been implicated in aberrant activities of neural circuits that play key roles in the pathogenesis of neurological and psychiatric diseases. Dopaminergic neurons are vulnerable to environmental insults. The neurotoxin methylmercury (MeHg) produces dopaminergic neuron damage in rodent as well as in Caenorhabditis elegans (C. elegans) models. Previous studies have demonstrated the utility of C. elegans as an alternative and complementary experimental model in dissecting out mechanism of MeHg-induced dopaminergic neurodegeneration. However, a sensitive pathological change that marks early events in neurodegeneration induced by environmental level of MeHg, is still lacking. By establishing a chronic exposure C. elegans model, for the first time, we have shown the propensity of MeHg (5 μM, 10 days) to induce bright puncta of dat-1::mCherry aggreagtes in the dendrites of cephalic (2 CEPs) dopaminergic neurons in a dose- and time-dependent manner, while these changes were not found in other dopaminergic neurons: anterior deirids (2 ADEs) and posterior deirids (2 PDEs), cholinergic neurons (2 AIYs) or glutamatergic neurons (2 PVDs). The bright puncta appear as an aggregation of mCherry proteins accumulating in dendrites. Further staining shows that the puncta were not inclusions in lysosome, or amyloid protein aggregates. In addition, features of the puncta including enlarged sphere shape (0.5-2 μm diameters), bright and accompanying with the shrinkage of the dendrite suggest that the puncta are likely composed of homologous mCherry molecules packaged at the dendritic site for exportation. Moreover, in the glutathione S-transferase 4 (gst-4) transcriptional reporter strain and RT-PCR assay, the expression levels of gst-4 and tubulins (tba-1 and tba-2) genes were not significantly modified under this chronic exposure paradigm, but gst-4 did show significant changes in an one day exposure paradigm. Collectively, these results suggest that CEP dopaminergic neurons are a sensitive target of MeHg, and the current exposure paradigm could be used as a model to investigate mechanism of dopaminergic neurotoxicity.
Keywords: C. elegansdopamine; Methylmercury; Toxicity; puncta formation.
Copyright © 2020 Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors report no declarations of interest.
Figures
References
-
- Angelo G, Van Gilst MR, 2009. Starvation protects germline stem cells and extends reproductive longevity in C. elegans. Science 326(5955), 954–958. - PubMed
-
- Arvidson B, 1987. Retrograde axonal transport of mercury. Exp Neurol 98(1), 198–203. - PubMed
-
- Arvidson B, 1990. Accumulation of mercury in brainstem nuclei of mice after retrograde axonal transport. Acta Neurol Scand 82(4), 234–237. - PubMed
-
- Arvidson B, 1992. Accumulation of Inorganic Mercury in Lower Motoneurons of Mice. Neurotoxicology 13(1), 277–280. - PubMed
-
- Arvidson B, Arvidsson J, 1990. Retrograde axonal transport of mercury in primary sensory neurons innervating the tooth pulp in the rat. Neurosci Lett 115(1), 29–32. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
