

Modulating Tumor-Associated Macrophage Polarization by Synthetic and Natural PPARγ Ligands as a Potential Target in Breast Cancer
- PMID: 31936729
- PMCID: PMC7017381
- DOI: 10.3390/cells9010174
Modulating Tumor-Associated Macrophage Polarization by Synthetic and Natural PPARγ Ligands as a Potential Target in Breast Cancer
Abstract
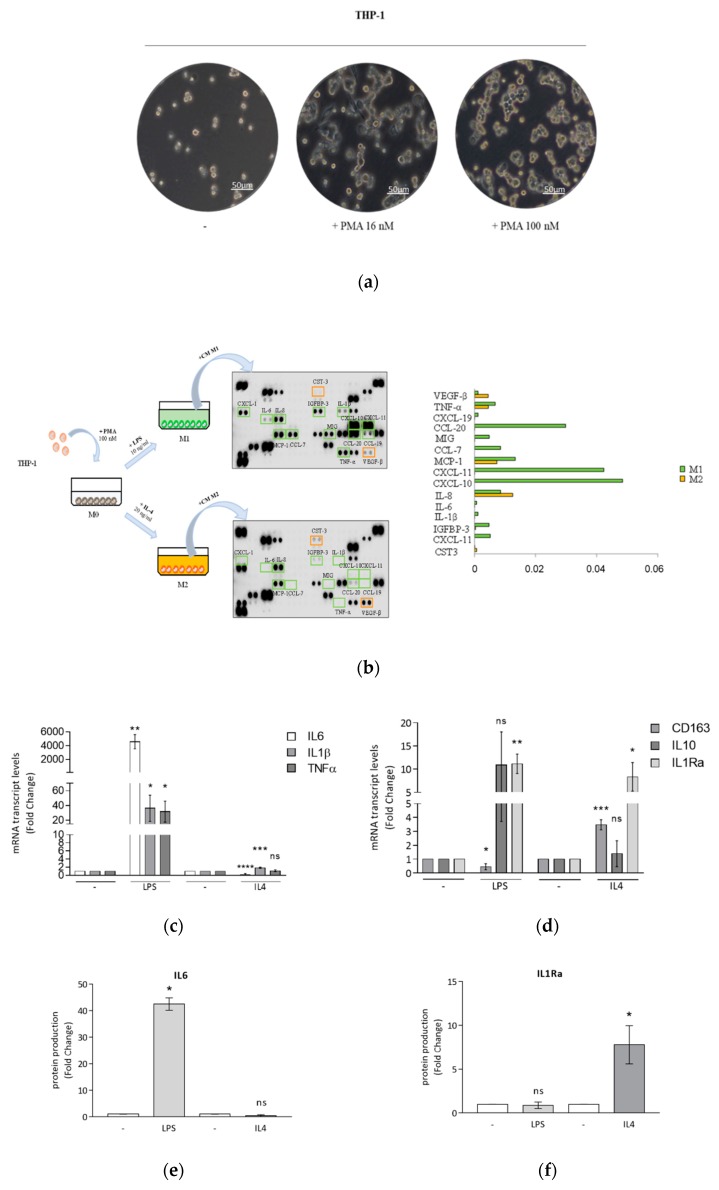
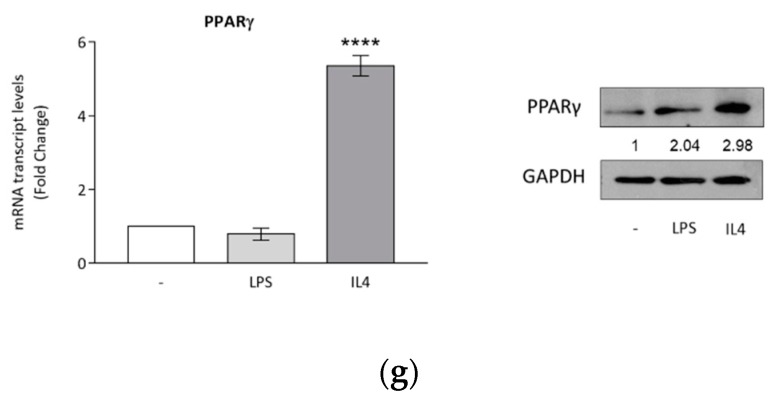
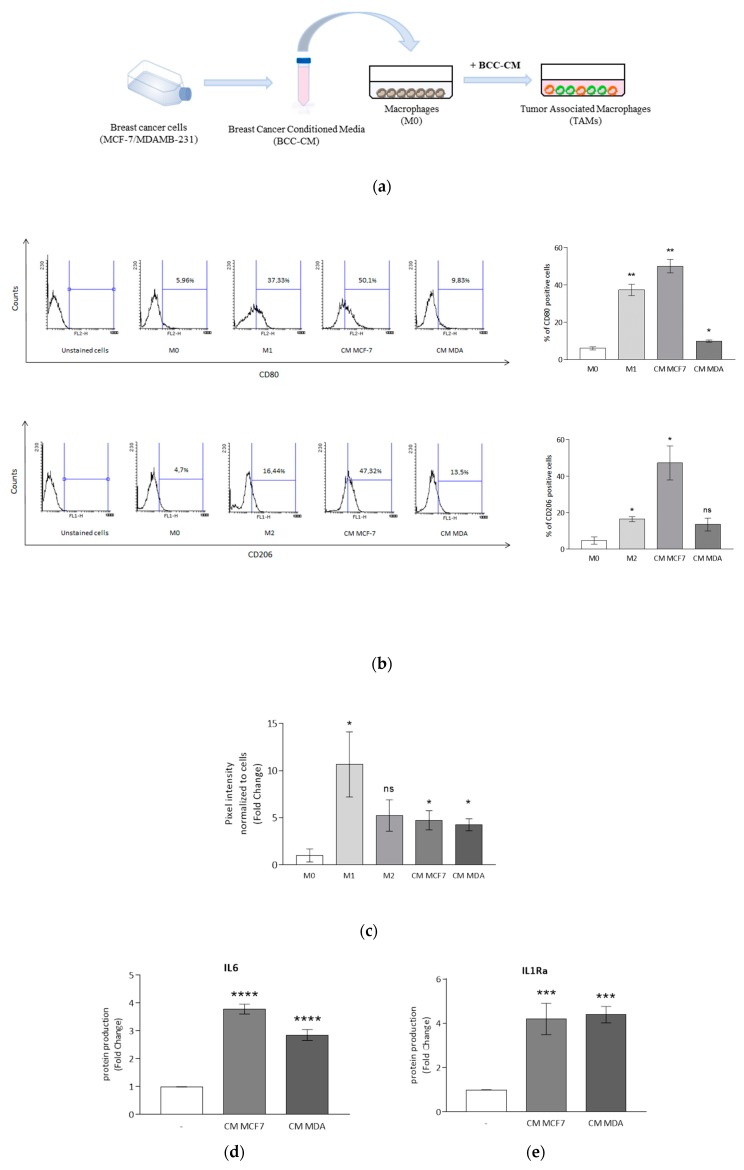
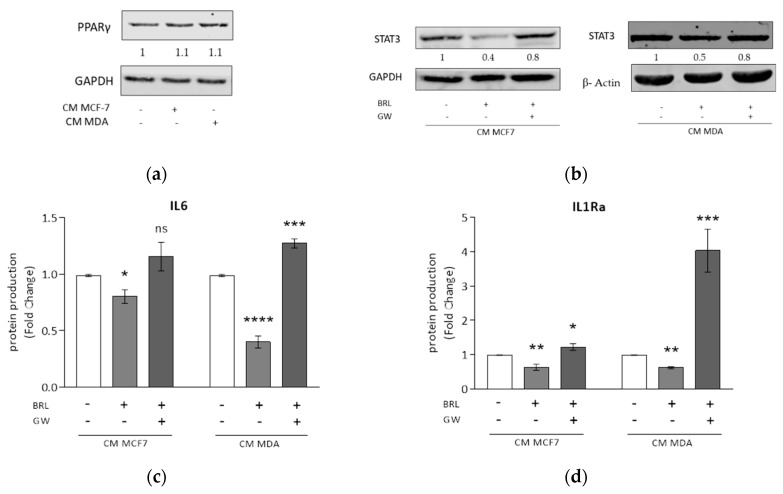
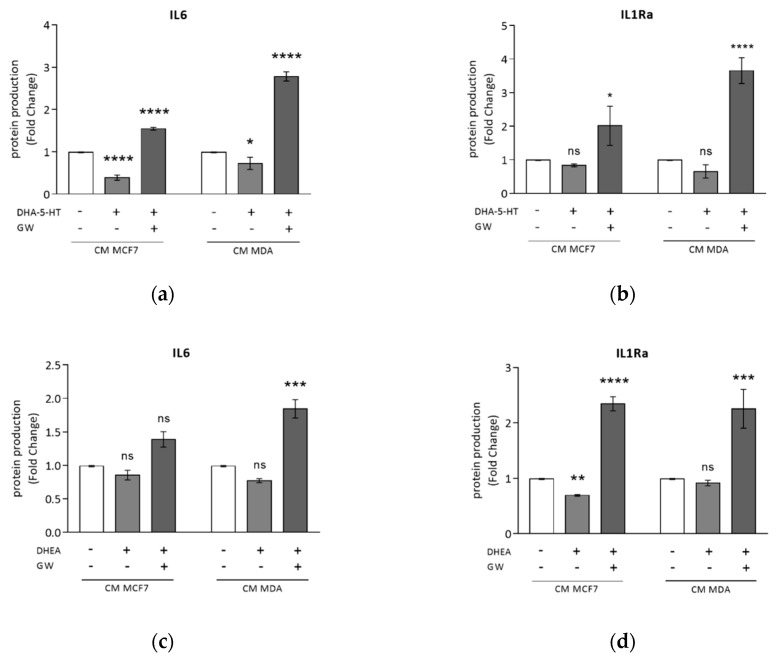
Activation of peroxisome proliferator-activated receptor gamma (PPARγ) elicits anti-proliferative effects on different tumor cells, including those derived from breast cancer. PPARγ is also expressed in several cells of the breast tumor microenvironment, among which tumor associated macrophages (TAMs) play a pivotal role in tumor progression and metastasis. We explored the ability of synthetic and natural PPARγ ligands to modulate TAM polarization. The ligands included rosiglitazone (BRL-49653), and two docosahexaenoic acid (DHA) conjugates, N-docosahexaenoyl ethanolamine (DHEA) and N-docosahexaenoyl serotonin (DHA-5-HT). Human THP-1 monocytic cells were differentiated into M0, M1 and M2 macrophages that were characterized by qRT-PCR, ELISA and western blotting. A TAM-like phenotypic state was generated by adding two different breast cancer cell conditioned media (BCC-CM) to the cultures. Macrophages exposed to BCC-CM concomitantly exhibited M1 and M2 phenotypes. Interestingly, rosiglitazone, DHEA and DHA-5-HT attenuated cytokine secretion by TAMs, and this effect was reversed by the PPARγ antagonist GW9662. Given the key role played by PPARγ in the crosstalk between cancer cells and TAMs in tumor progression, its activation via endogenous or synthetic ligands may lead to novel strategies that target both epithelial neoplastic cells and the tumor microenvironment.
Keywords: breast tumor microenvironment; docosahexaenoyl ethanolamine (DHEA); docosahexaenoyl serotonin (DHA-5-HT); n-3 polyunsaturated fatty acids; peroxisome proliferator-activated receptor gamma; rosiglitazone; tumor associated macrophages.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
M2 polarization of tumor-associated macrophages is dependent on integrin β3 via peroxisome proliferator-activated receptor-γ up-regulation in breast cancer.Immunology. 2020 Aug;160(4):345-356. doi: 10.1111/imm.13196. Epub 2020 Apr 29. Immunology. 2020. PMID: 32311768 Free PMC article.
-
Omega-3 PUFA ethanolamides DHEA and EPEA induce autophagy through PPARγ activation in MCF-7 breast cancer cells.J Cell Physiol. 2013 Jun;228(6):1314-22. doi: 10.1002/jcp.24288. J Cell Physiol. 2013. PMID: 23168911
-
Docosahexaenoic acid induces M2 macrophage polarization through peroxisome proliferator-activated receptor γ activation.Life Sci. 2015 Jan 1;120:39-47. doi: 10.1016/j.lfs.2014.10.014. Epub 2014 Oct 31. Life Sci. 2015. PMID: 25445227
-
Natural and Synthetic PPARγ Ligands in Tumor Microenvironment: A New Potential Strategy against Breast Cancer.Int J Mol Sci. 2020 Dec 19;21(24):9721. doi: 10.3390/ijms21249721. Int J Mol Sci. 2020. PMID: 33352766 Free PMC article. Review.
-
Docosahexaenoic acid attenuates LPS-stimulated inflammatory response by regulating the PPARγ/NF-κB pathways in primary bovine mammary epithelial cells.Res Vet Sci. 2017 Jun;112:7-12. doi: 10.1016/j.rvsc.2016.12.011. Epub 2017 Jan 5. Res Vet Sci. 2017. PMID: 28095338 Review.
Cited by
-
PPARgamma: A Potential Intrinsic and Extrinsic Molecular Target for Breast Cancer Therapy.Biomedicines. 2021 May 13;9(5):543. doi: 10.3390/biomedicines9050543. Biomedicines. 2021. PMID: 34067944 Free PMC article.
-
CEBP-β and PLK1 as Potential Mediators of the Breast Cancer/Obesity Crosstalk: In Vitro and In Silico Analyses.Nutrients. 2023 Jun 22;15(13):2839. doi: 10.3390/nu15132839. Nutrients. 2023. PMID: 37447165 Free PMC article.
-
Peroxisome Proliferator-Activated Receptors and the Hallmarks of Cancer.Cells. 2022 Aug 5;11(15):2432. doi: 10.3390/cells11152432. Cells. 2022. PMID: 35954274 Free PMC article. Review.
-
TRAIL promotes the polarization of human macrophages toward a proinflammatory M1 phenotype and is associated with increased survival in cancer patients with high tumor macrophage content.Front Immunol. 2023 Sep 21;14:1209249. doi: 10.3389/fimmu.2023.1209249. eCollection 2023. Front Immunol. 2023. PMID: 37809073 Free PMC article.
-
Effects of Mediterranean Diet on Chronic Degenerative Diseases and Human Healthy Lifestyle.Nutrients. 2025 Apr 1;17(7):1231. doi: 10.3390/nu17071231. Nutrients. 2025. PMID: 40218989 Free PMC article.
References
-
- Willett W.C. Diet and cancer: One view at the start of the millennium. Cancer Epidemiol. Biomark. Prev. 2001;10:3–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
