

Optogenetic activation of parvalbumin and somatostatin interneurons selectively restores theta-nested gamma oscillations and oscillation-induced spike timing-dependent long-term potentiation impaired by amyloid β oligomers
- PMID: 31937327
- PMCID: PMC6961381
- DOI: 10.1186/s12915-019-0732-7
Optogenetic activation of parvalbumin and somatostatin interneurons selectively restores theta-nested gamma oscillations and oscillation-induced spike timing-dependent long-term potentiation impaired by amyloid β oligomers
Abstract
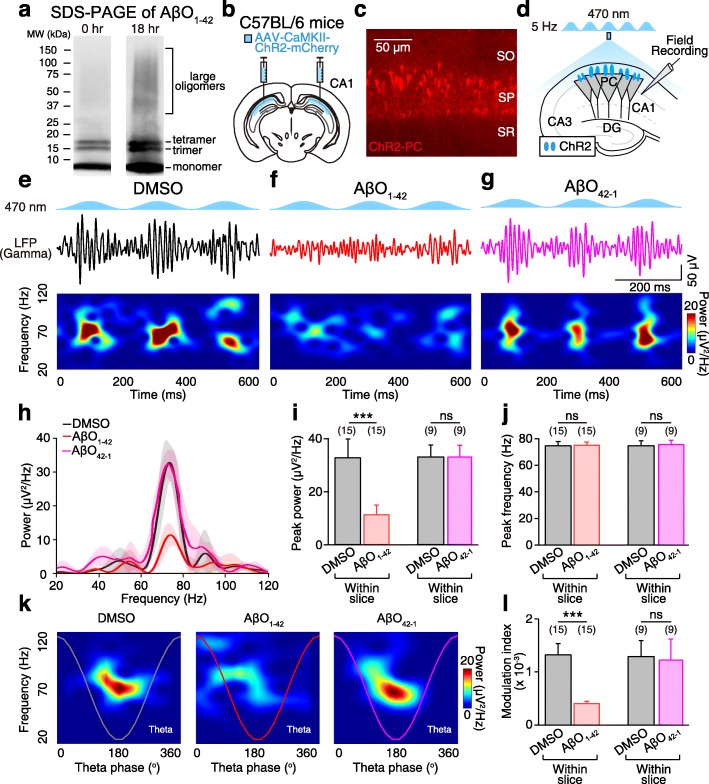
Background: Abnormal accumulation of amyloid β1-42 oligomers (AβO1-42), a hallmark of Alzheimer's disease, impairs hippocampal theta-nested gamma oscillations and long-term potentiation (LTP) that are believed to underlie learning and memory. Parvalbumin-positive (PV) and somatostatin-positive (SST) interneurons are critically involved in theta-nested gamma oscillogenesis and LTP induction. However, how AβO1-42 affects PV and SST interneuron circuits is unclear. Through optogenetic manipulation of PV and SST interneurons and computational modeling of the hippocampal neural circuits, we dissected the contributions of PV and SST interneuron circuit dysfunctions on AβO1-42-induced impairments of hippocampal theta-nested gamma oscillations and oscillation-induced LTP.
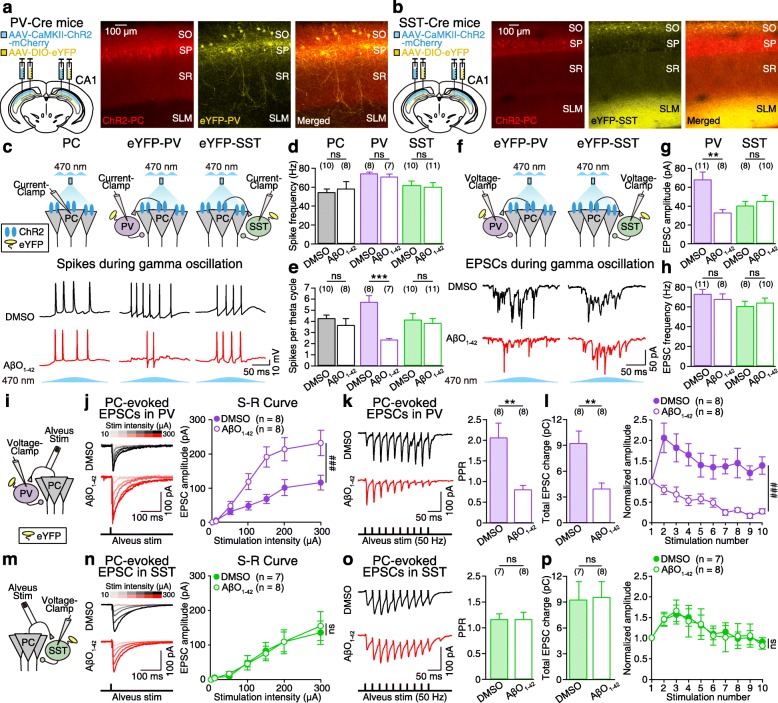
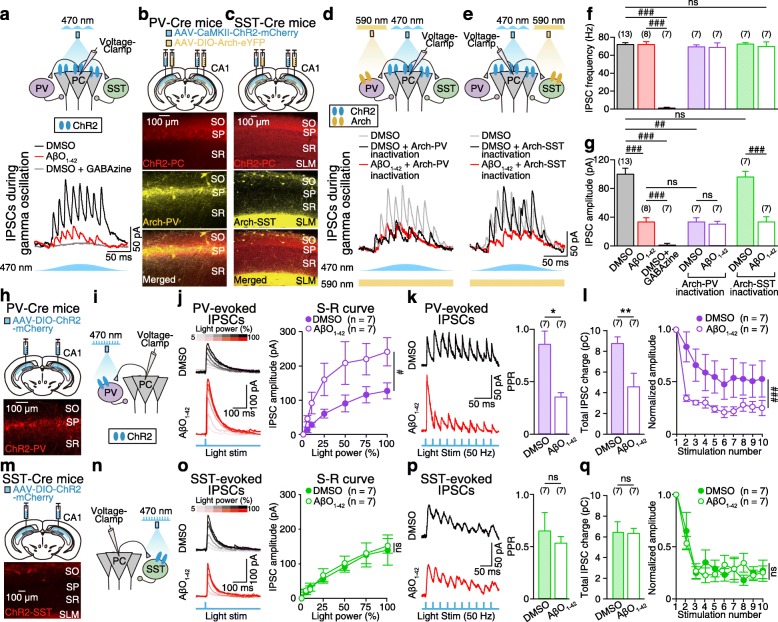
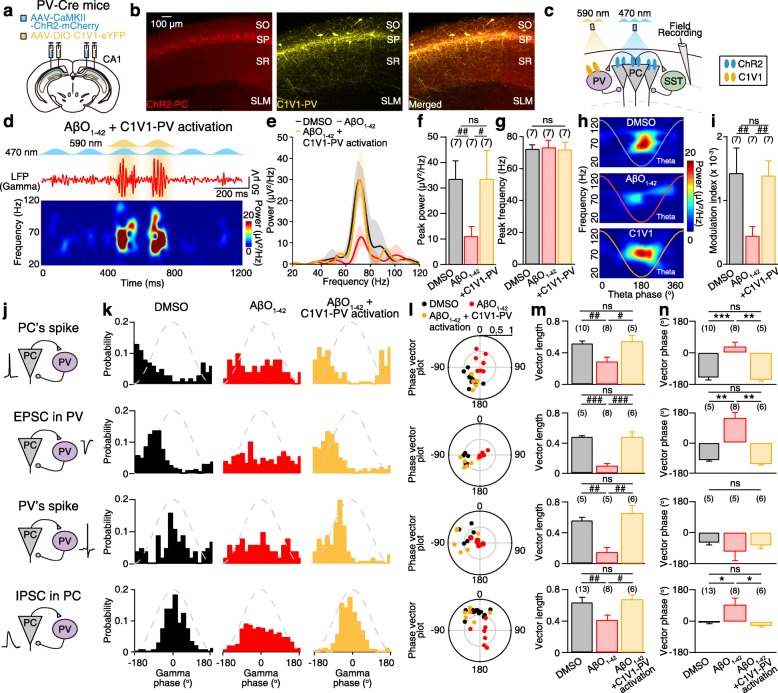
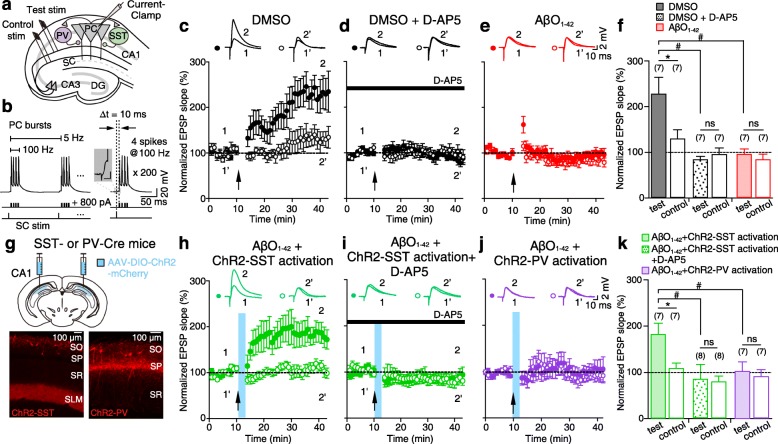
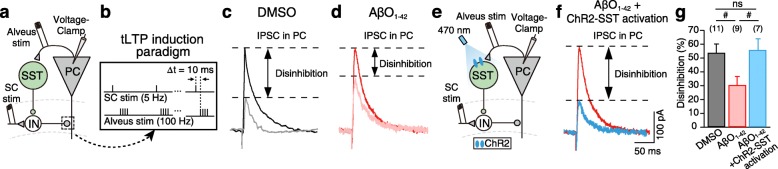
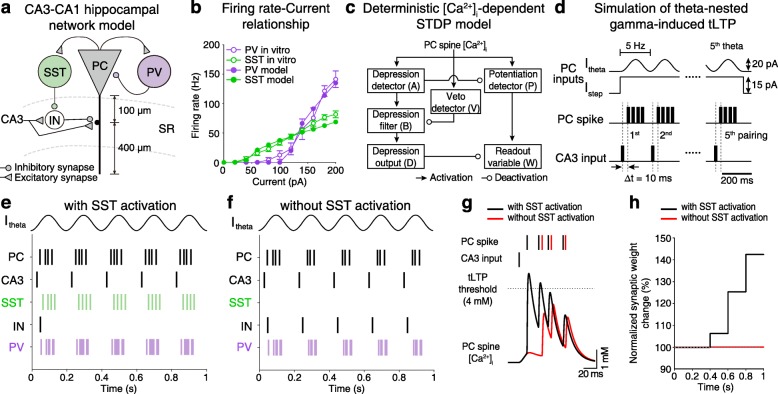
Results: Targeted whole-cell patch-clamp recordings and optogenetic manipulations of PV and SST interneurons during in vivo-like, optogenetically induced theta-nested gamma oscillations in vitro revealed that AβO1-42 causes synapse-specific dysfunction in PV and SST interneurons. AβO1-42 selectively disrupted CA1 pyramidal cells (PC)-to-PV interneuron and PV-to-PC synapses to impair theta-nested gamma oscillogenesis. In contrast, while having no effect on PC-to-SST or SST-to-PC synapses, AβO1-42 selectively disrupted SST interneuron-mediated disinhibition to CA1 PC to impair theta-nested gamma oscillation-induced spike timing-dependent LTP (tLTP). Such AβO1-42-induced impairments of gamma oscillogenesis and oscillation-induced tLTP were fully restored by optogenetic activation of PV and SST interneurons, respectively, further supporting synapse-specific dysfunctions in PV and SST interneurons. Finally, computational modeling of hippocampal neural circuits including CA1 PC, PV, and SST interneurons confirmed the experimental observations and further revealed distinct functional roles of PV and SST interneurons in theta-nested gamma oscillations and tLTP induction.
Conclusions: Our results reveal that AβO1-42 causes synapse-specific dysfunctions in PV and SST interneurons and that optogenetic modulations of these interneurons present potential therapeutic targets for restoring hippocampal network oscillations and synaptic plasticity impairments in Alzheimer's disease.
Keywords: Alzheimer’s disease; Amyloid beta oligomers; Hippocampus; Optogenetics; Parvalbumin interneuron; Somatostatin interneuron; Spike timing-dependent long-term potentiation; Synapse-specific dysfunction; Theta-nested gamma oscillations.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Dissociation of somatostatin and parvalbumin interneurons circuit dysfunctions underlying hippocampal theta and gamma oscillations impaired by amyloid β oligomers in vivo.Brain Struct Funct. 2020 Apr;225(3):935-954. doi: 10.1007/s00429-020-02044-3. Epub 2020 Feb 27. Brain Struct Funct. 2020. PMID: 32107637 Free PMC article.
-
Parvalbumin and Somatostatin Interneurons Contribute to the Generation of Hippocampal Gamma Oscillations.J Neurosci. 2020 Sep 30;40(40):7668-7687. doi: 10.1523/JNEUROSCI.0261-20.2020. Epub 2020 Aug 28. J Neurosci. 2020. PMID: 32859716 Free PMC article.
-
Excitatory Inputs Determine Phase-Locking Strength and Spike-Timing of CA1 Stratum Oriens/Alveus Parvalbumin and Somatostatin Interneurons during Intrinsically Generated Hippocampal Theta Rhythm.J Neurosci. 2016 Jun 22;36(25):6605-22. doi: 10.1523/JNEUROSCI.3951-13.2016. J Neurosci. 2016. PMID: 27335395 Free PMC article.
-
A Role for Somatostatin-Positive Interneurons in Neuro-Oscillatory and Information Processing Deficits in Schizophrenia.Schizophr Bull. 2021 Aug 21;47(5):1385-1398. doi: 10.1093/schbul/sbaa184. Schizophr Bull. 2021. PMID: 33370434 Free PMC article. Review.
-
Parvalbumin Interneuron Dysfunction in Neurological Disorders: Focus on Epilepsy and Alzheimer's Disease.Int J Mol Sci. 2024 May 19;25(10):5549. doi: 10.3390/ijms25105549. Int J Mol Sci. 2024. PMID: 38791587 Free PMC article. Review.
Cited by
-
Parvalbumin interneuron-mediated neural disruption in an animal model of postintensive care syndrome: prevention by fluoxetine.Aging (Albany NY). 2021 Feb 22;13(6):8720-8736. doi: 10.18632/aging.202684. Epub 2021 Feb 22. Aging (Albany NY). 2021. PMID: 33619236 Free PMC article.
-
Mystery of gamma wave stimulation in brain disorders.Mol Neurodegener. 2024 Dec 18;19(1):96. doi: 10.1186/s13024-024-00785-x. Mol Neurodegener. 2024. PMID: 39695746 Free PMC article. Review.
-
Resilient Hippocampal Gamma Rhythmogenesis and Parvalbumin-Expressing Interneuron Function Before and After Plaque Burden in 5xFAD Alzheimer's Disease Model.Front Synaptic Neurosci. 2022 May 11;14:857608. doi: 10.3389/fnsyn.2022.857608. eCollection 2022. Front Synaptic Neurosci. 2022. PMID: 35645763 Free PMC article.
-
Cacao consumption improves passive avoidance memory impairment in a rat model of Alzheimer's disease: the role of hippocampal synaptic plasticity and oxidative stress.Front Pharmacol. 2024 May 2;15:1379264. doi: 10.3389/fphar.2024.1379264. eCollection 2024. Front Pharmacol. 2024. PMID: 38756381 Free PMC article.
-
The Pathophysiological Underpinnings of Gamma-Band Alterations in Psychiatric Disorders.Life (Basel). 2024 Apr 30;14(5):578. doi: 10.3390/life14050578. Life (Basel). 2024. PMID: 38792599 Free PMC article. Review.
References
-
- Lambert MP, Barlow AK, Chromy BA, Edwards C, Freed R, Liosatos M, Morgan TE, Rozovsky I, Trommer B, Viola KL, et al. Diffusible, nonfibrillar ligands derived from Abeta1-42 are potent central nervous system neurotoxins. Proc Natl Acad Sci U S A. 1998;95(11):6448–6453. doi: 10.1073/pnas.95.11.6448. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
