

Structure and activation mechanism of the BBSome membrane protein trafficking complex
- PMID: 31939736
- PMCID: PMC7018513
- DOI: 10.7554/eLife.53322
Structure and activation mechanism of the BBSome membrane protein trafficking complex
Abstract
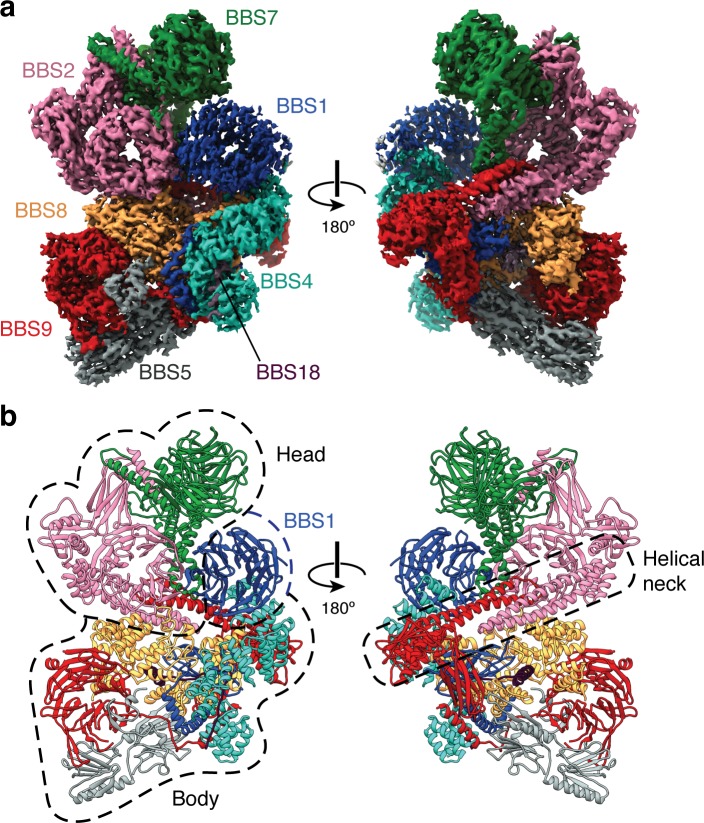
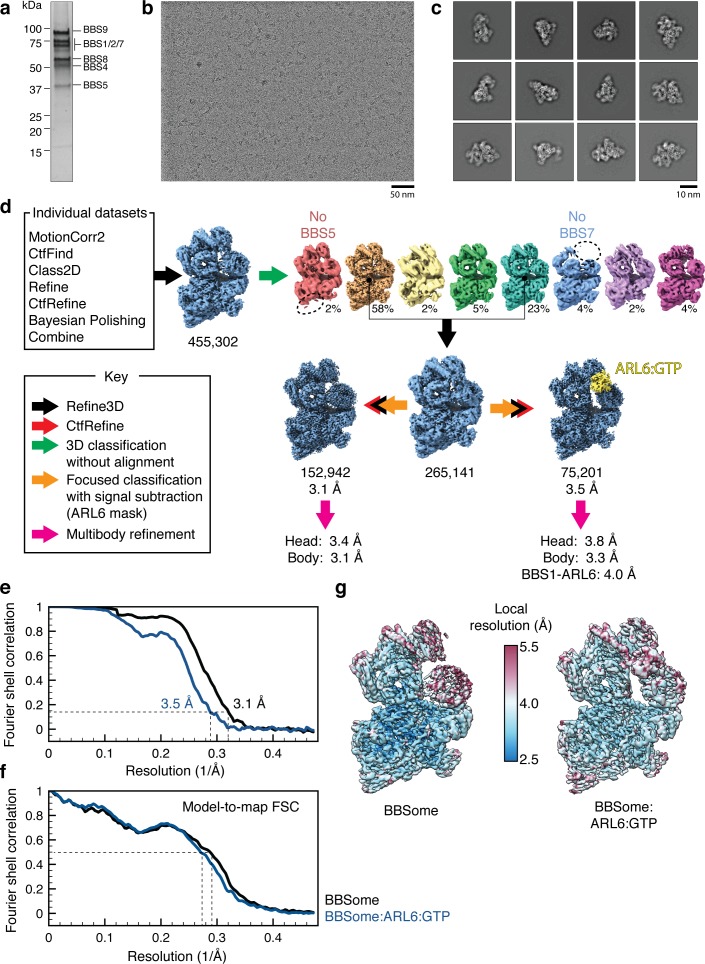
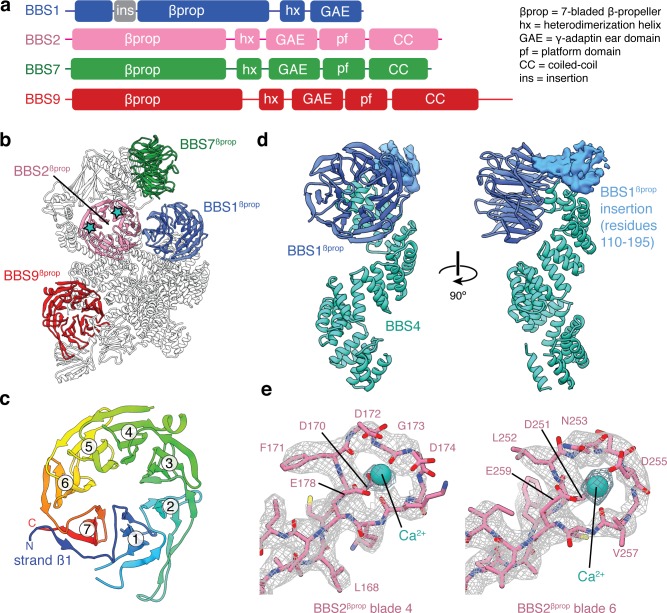
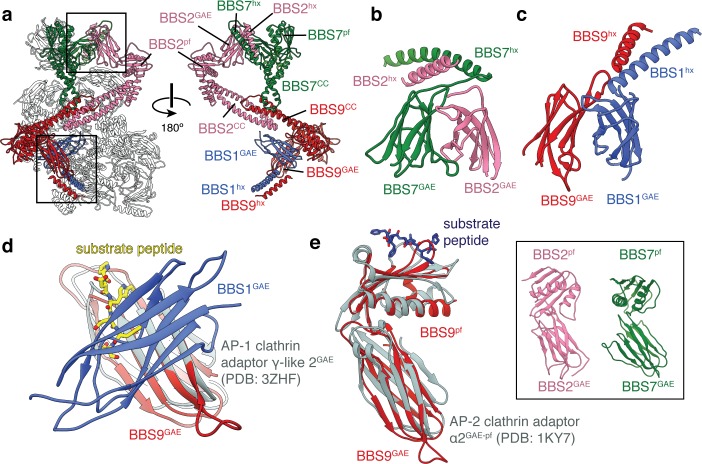
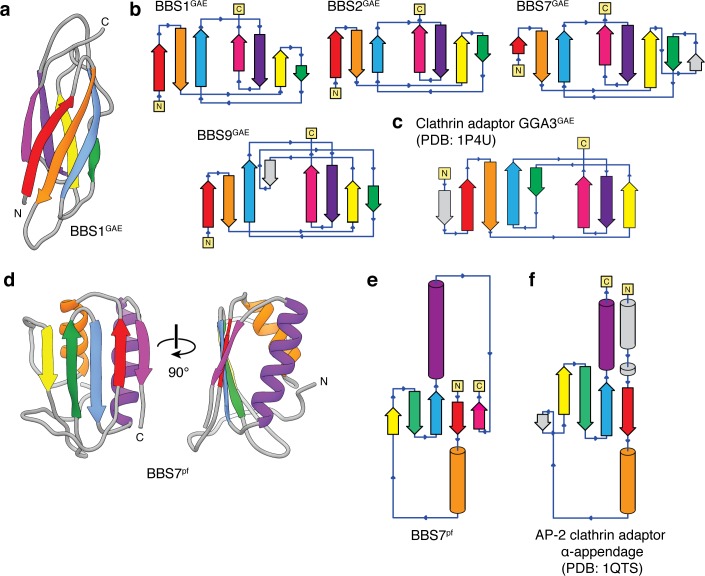
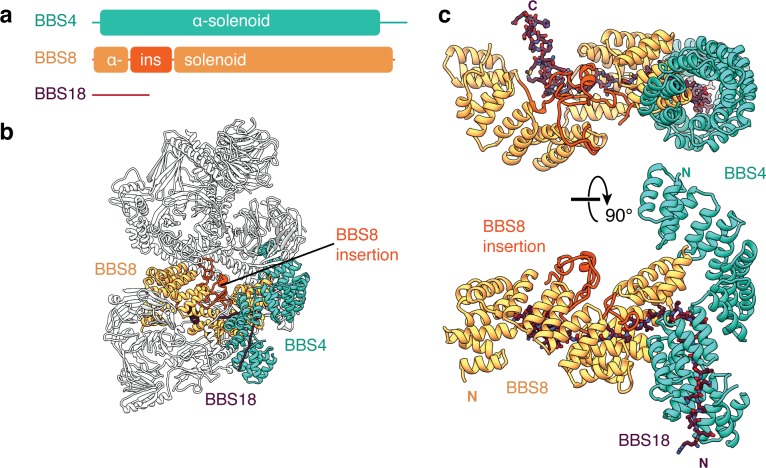
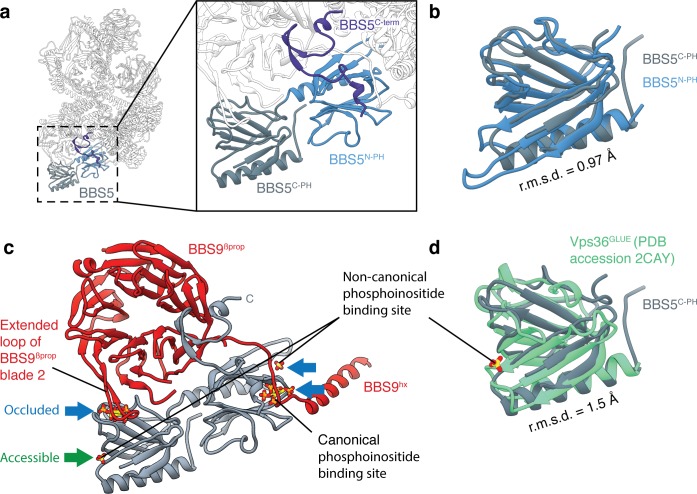
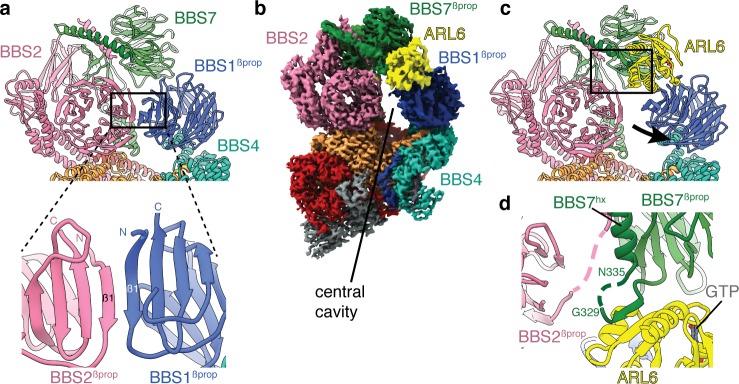
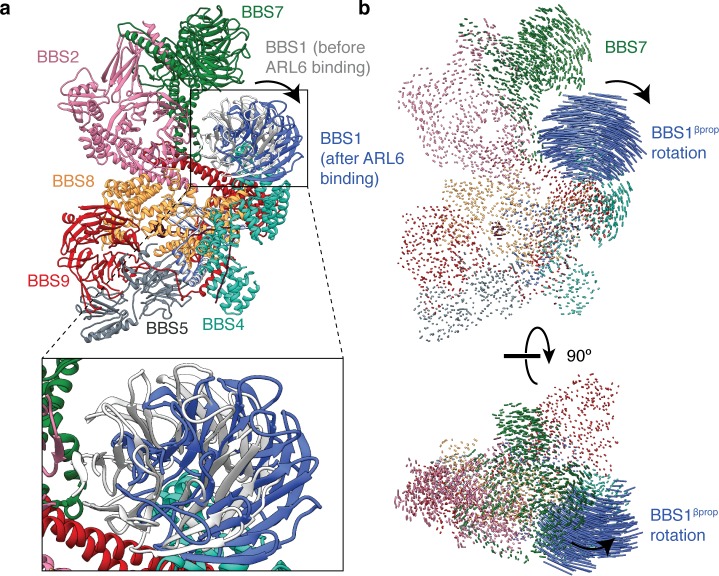
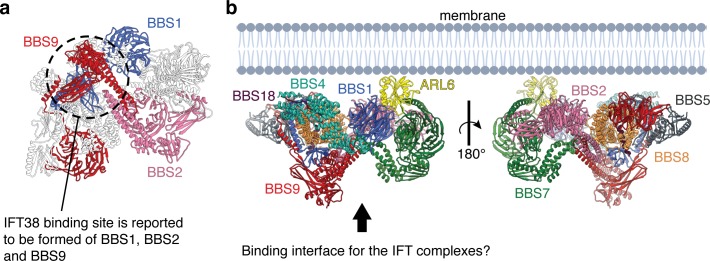
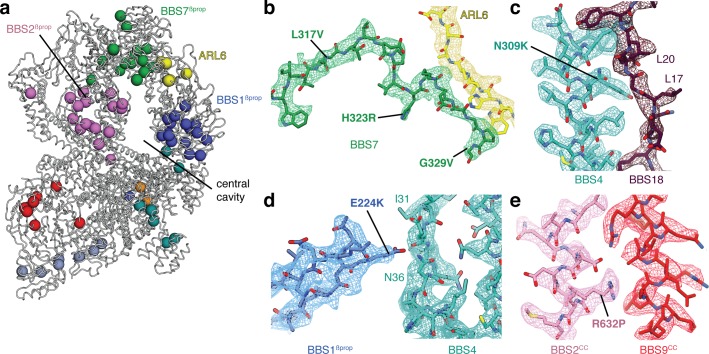
Bardet-Biedl syndrome (BBS) is a currently incurable ciliopathy caused by the failure to correctly establish or maintain cilia-dependent signaling pathways. Eight proteins associated with BBS assemble into the BBSome, a key regulator of the ciliary membrane proteome. We report the electron cryomicroscopy (cryo-EM) structures of the native bovine BBSome in inactive and active states at 3.1 and 3.5 Å resolution, respectively. In the active state, the BBSome is bound to an Arf-family GTPase (ARL6/BBS3) that recruits the BBSome to ciliary membranes. ARL6 recognizes a composite binding site formed by BBS1 and BBS7 that is occluded in the inactive state. Activation requires an unexpected swiveling of the β-propeller domain of BBS1, the subunit most frequently implicated in substrate recognition, which widens a central cavity of the BBSome. Structural mapping of disease-causing mutations suggests that pathogenesis results from folding defects and the disruption of autoinhibition and activation.
Keywords: BBSome; Bos taurus; cilia; cryo-EM; intraflagellar transport; molecular biophysics; protein trafficking; structural biology.
© 2020, Singh et al.
Conflict of interest statement
SS, MG, MY, AB No competing interests declared, FK is affiliated with Thermo Fisher Scientific. The author has no financial interests to declare
Figures

Comment in
-
Moving proteins along in the cilium.Elife. 2020 Feb 13;9:e55254. doi: 10.7554/eLife.55254. Elife. 2020. PMID: 32052743 Free PMC article.
References
-
- Abd-El-Barr MM, Sykoudis K, Andrabi S, Eichers ER, Pennesi ME, Tan PL, Wilson JH, Katsanis N, Lupski JR, Wu SM. Impaired photoreceptor protein transport and synaptic transmission in a mouse model of Bardet–Biedl syndrome. Vision Research. 2007;47:3394–3407. doi: 10.1016/j.visres.2007.09.016. - DOI - PMC - PubMed
-
- Aldahmesh MA, Li Y, Alhashem A, Anazi S, Alkuraya H, Hashem M, Awaji AA, Sogaty S, Alkharashi A, Alzahrani S, Al Hazzaa SA, Xiong Y, Kong S, Sun Z, Alkuraya FS. IFT27, encoding a small GTPase component of IFT particles, is mutated in a consanguineous family with Bardet-Biedl syndrome. Human Molecular Genetics. 2014;23:3307–3315. doi: 10.1093/hmg/ddu044. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
