

Mutation position is an important determinant for predicting cancer neoantigens
- PMID: 31940002
- PMCID: PMC7144530
- DOI: 10.1084/jem.20190179
Mutation position is an important determinant for predicting cancer neoantigens
Abstract
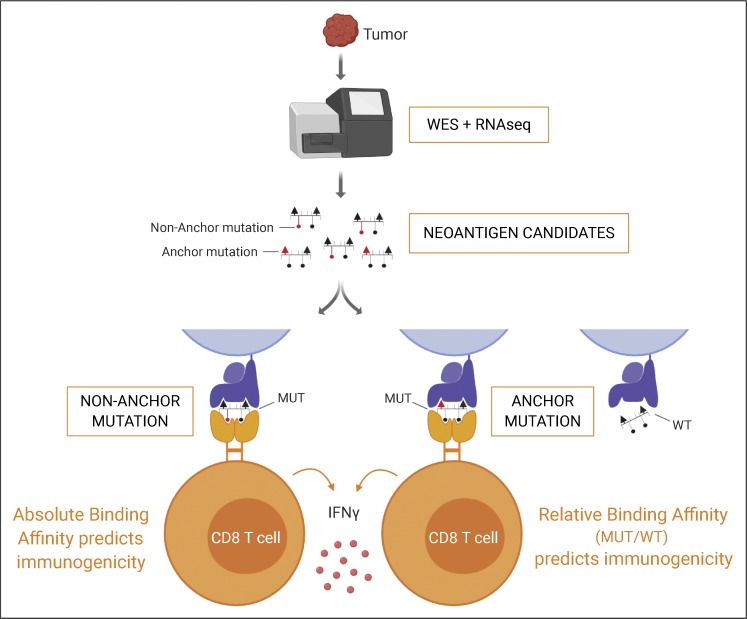
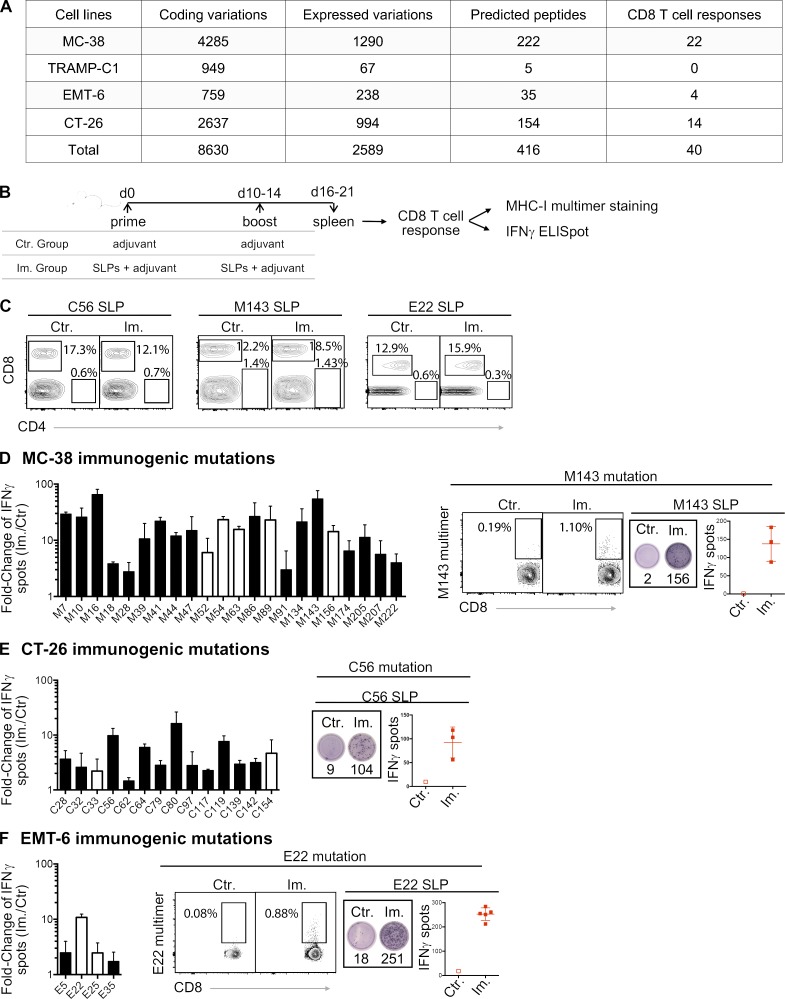
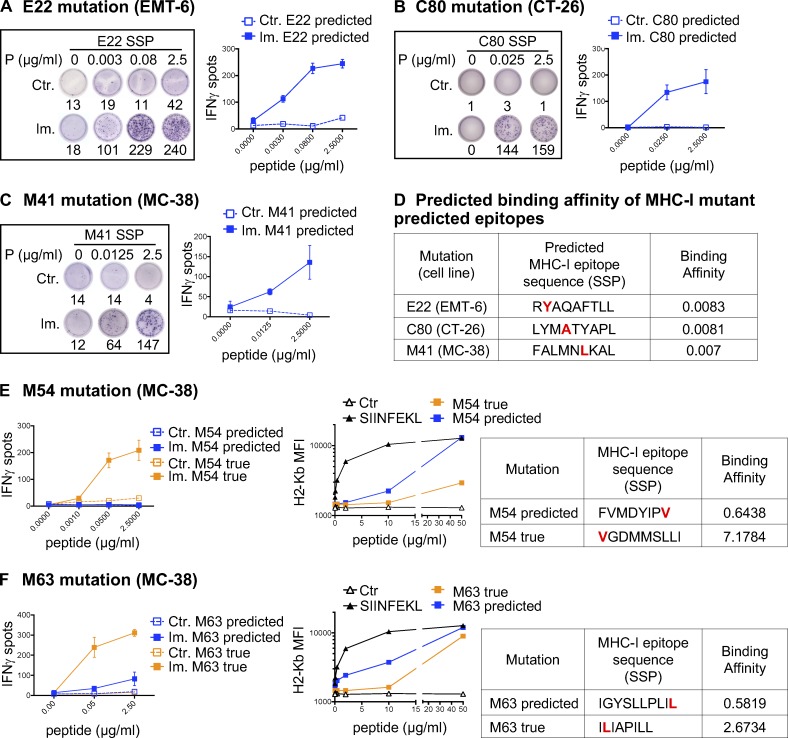
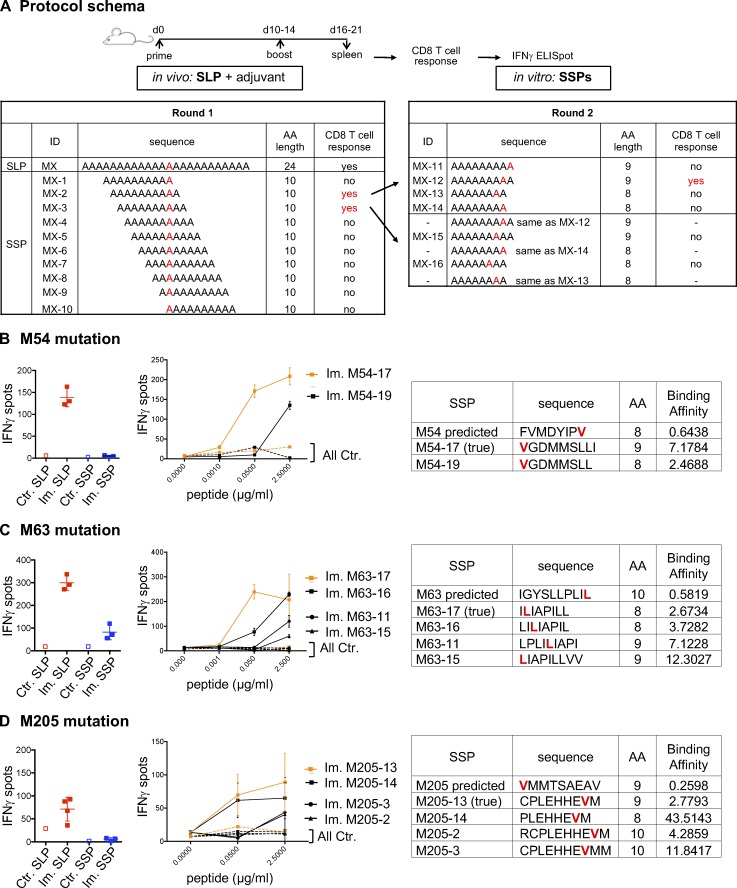
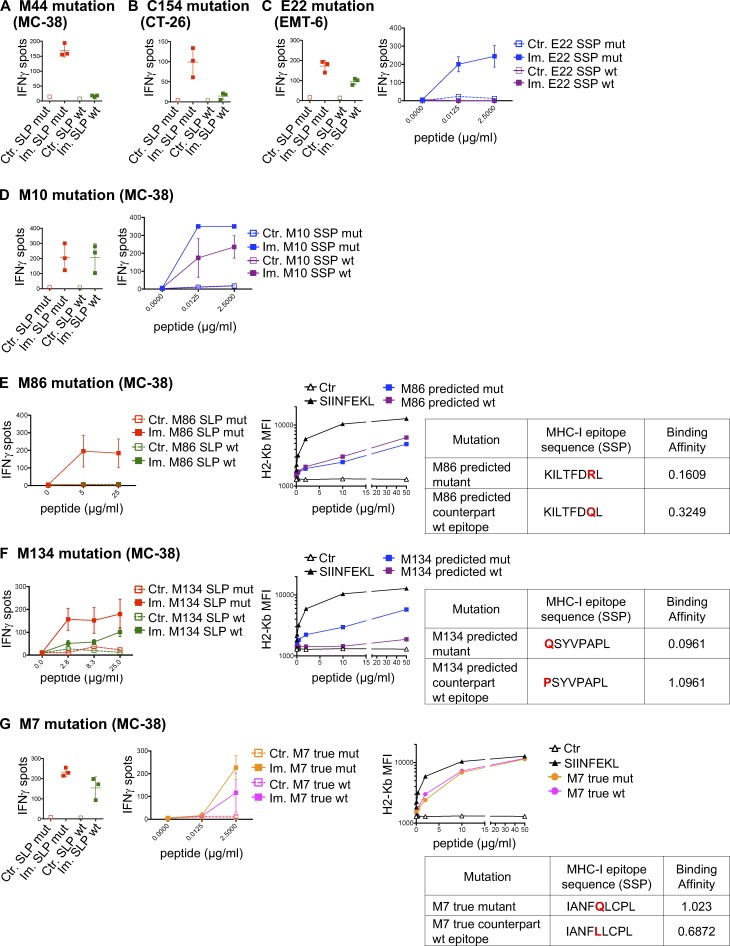
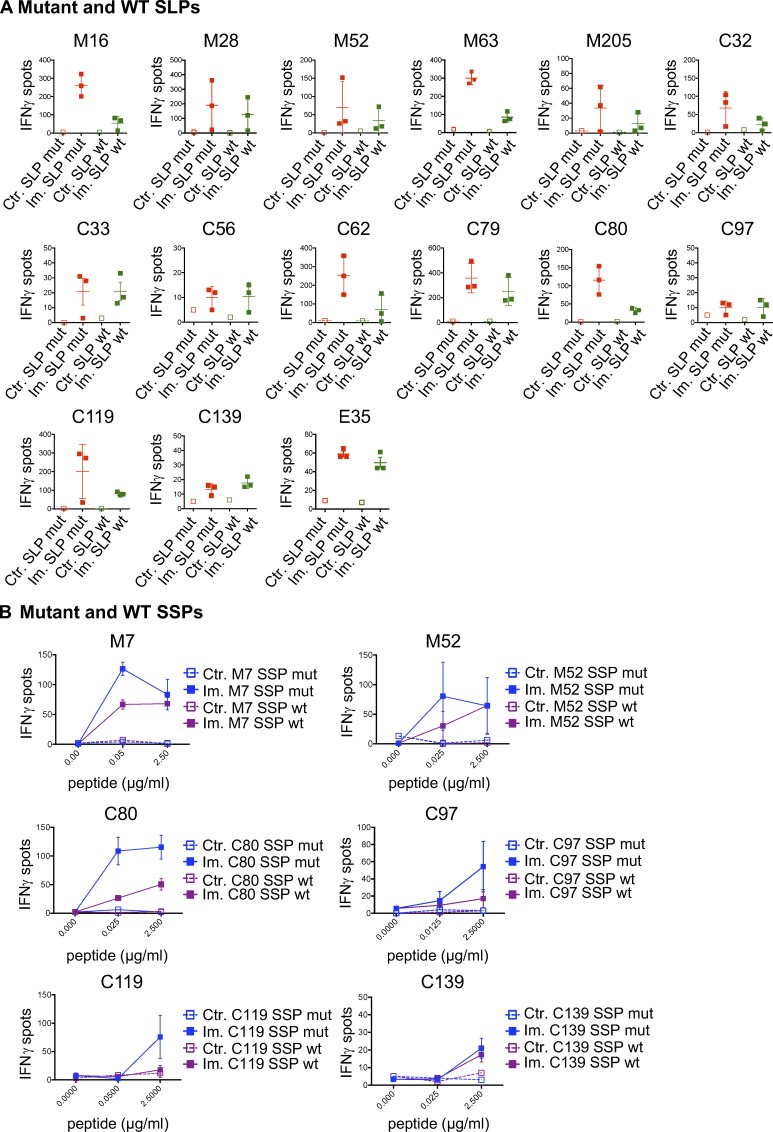
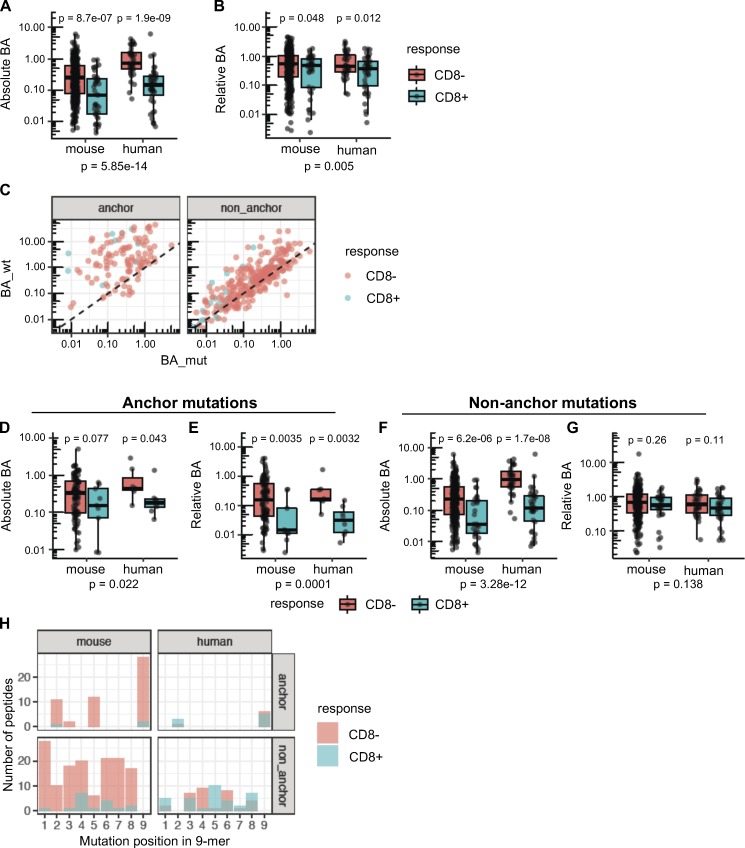
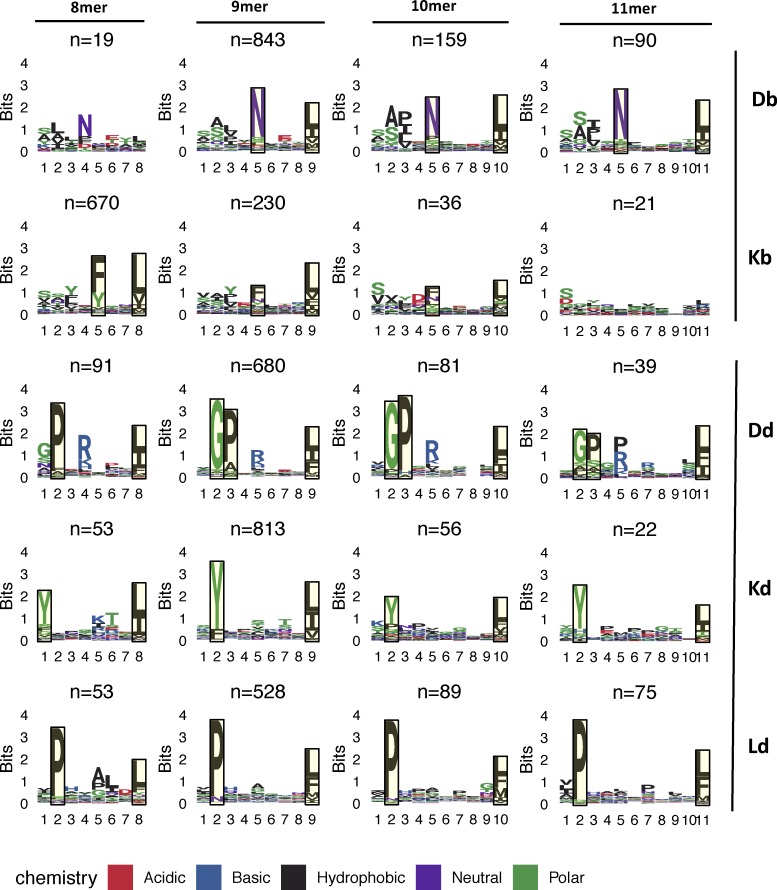
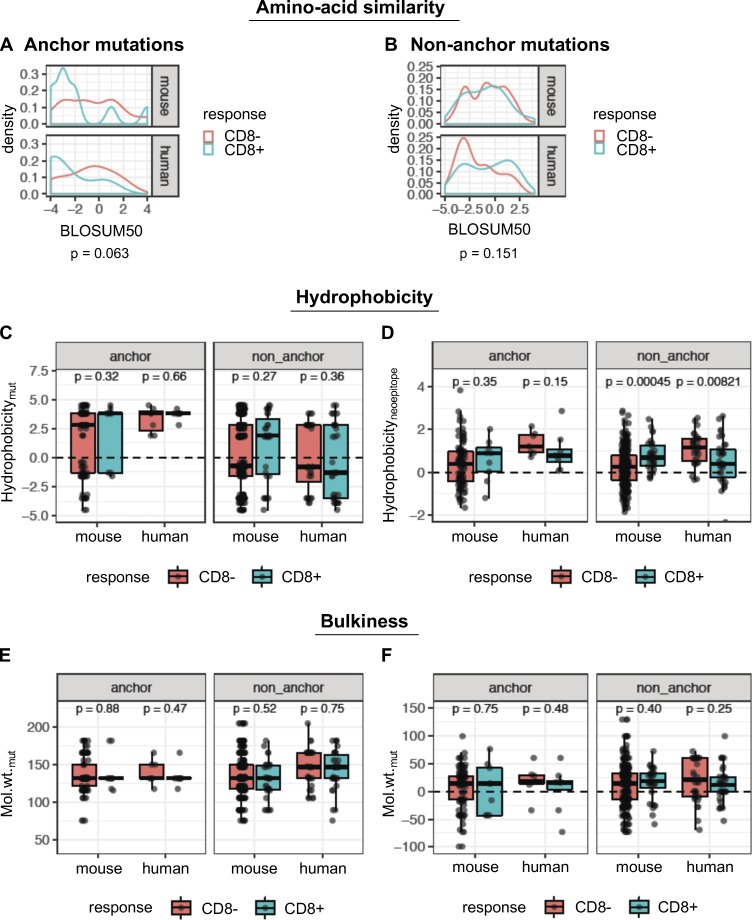
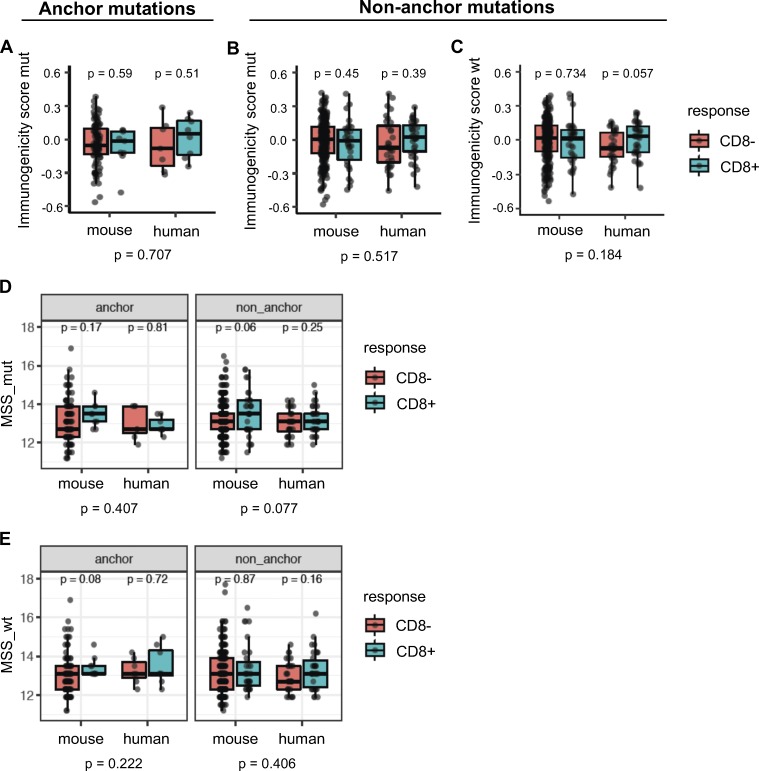
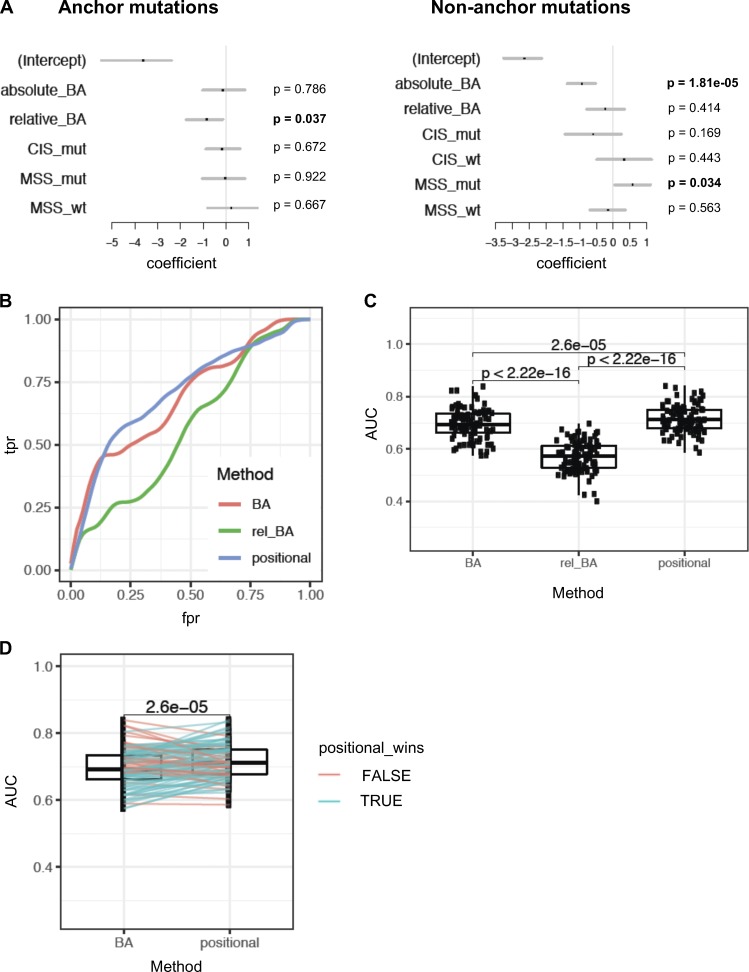
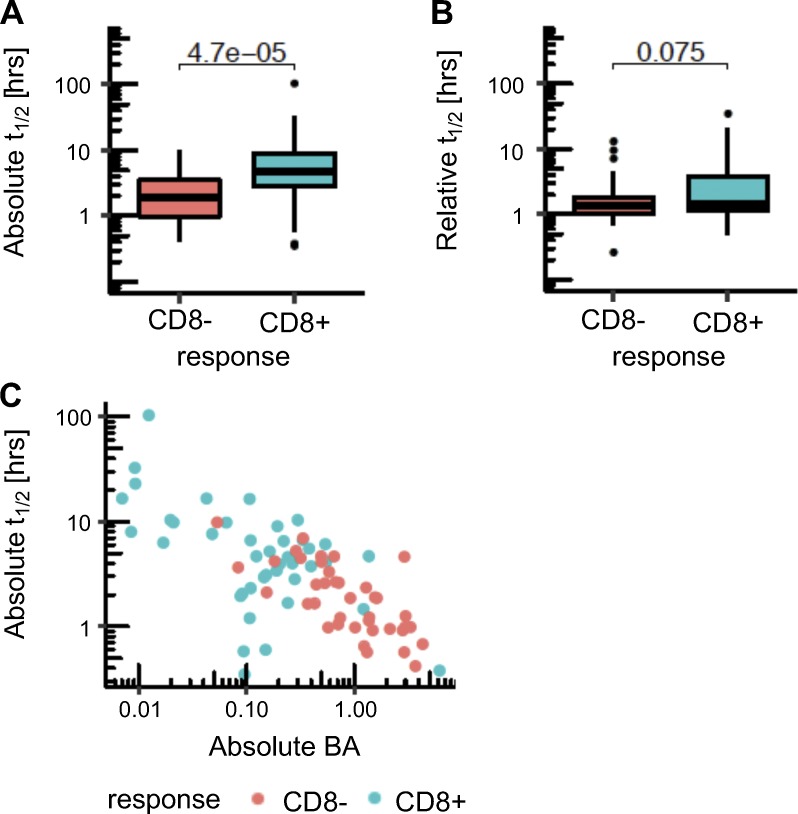
Tumor-specific mutations can generate neoantigens that drive CD8 T cell responses against cancer. Next-generation sequencing and computational methods have been successfully applied to identify mutations and predict neoantigens. However, only a small fraction of predicted neoantigens are immunogenic. Currently, predicted peptide binding affinity for MHC-I is often the major criterion for prioritizing neoantigens, although little progress has been made toward understanding the precise functional relationship between affinity and immunogenicity. We therefore systematically assessed the immunogenicity of peptides containing single amino acid mutations in mouse tumor models and divided them into two classes of immunogenic mutations. The first comprises mutations at a nonanchor residue, for which we find that the predicted absolute binding affinity is predictive of immunogenicity. The second involves mutations at an anchor residue; here, predicted relative affinity (compared with the WT counterpart) is a better predictor. Incorporating these features into an immunogenicity model significantly improves neoantigen ranking. Importantly, these properties of neoantigens are also predictive in human datasets, suggesting that they can be used to prioritize neoantigens for individualized neoantigen-specific immunotherapies.
© 2020 Capietto et al.
Conflict of interest statement
Disclosures: Dr. Jhunjhunwala, Dr. Lupardus, and Dr. Delamarre reported a patent to US 20160069895 A1 pending. Dr. Wong reported personal fees from Oncomed Pharmaceuticals, personal fees from Array BioPharma, and personal fees from Pfizer outside the submitted work. Dr. de la Cruz reported "other" from Genentech outside the submitted work. Dr. Mellman reported personal fees from Genentech and grants from Genentech during the conduct of the study. No other disclosures were reported.
Figures
References
-
- Abelin J.G., Keskin D.B., Sarkizova S., Hartigan C.R., Zhang W., Sidney J., Stevens J., Lane W., Zhang G.L., Eisenhaure T.M., et al. . 2017. Mass Spectrometry Profiling of HLA-Associated Peptidomes in Mono-allelic Cells Enables More Accurate Epitope Prediction. Immunity. 46:315–326. 10.1016/j.immuni.2017.02.007 - DOI - PMC - PubMed
-
- Bates D., Mächler M., Bolker B., and Walker S.. 2015. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67:1–48. 10.18637/jss.v067.i01 - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
