

Allergic inflammation is initiated by IL-33-dependent crosstalk between mast cells and basophils
- PMID: 31940364
- PMCID: PMC6961911
- DOI: 10.1371/journal.pone.0226701
Allergic inflammation is initiated by IL-33-dependent crosstalk between mast cells and basophils
Abstract
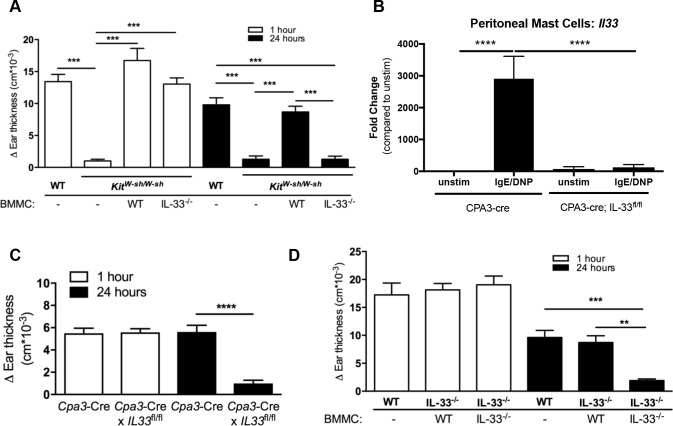
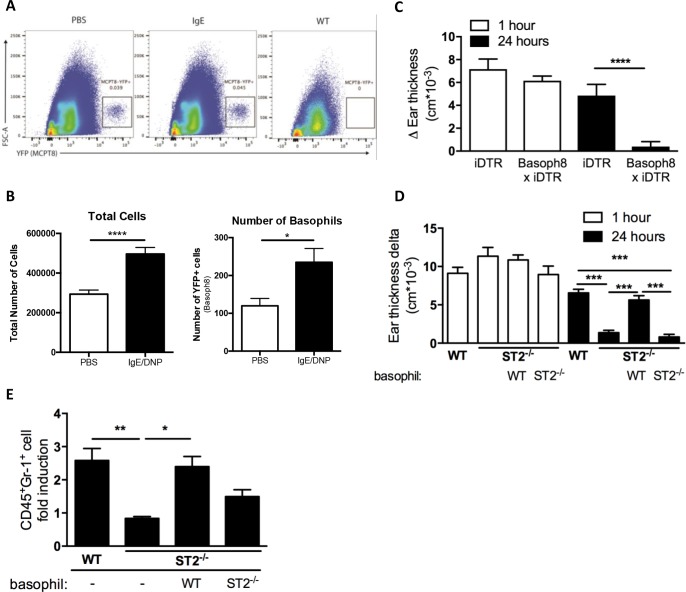
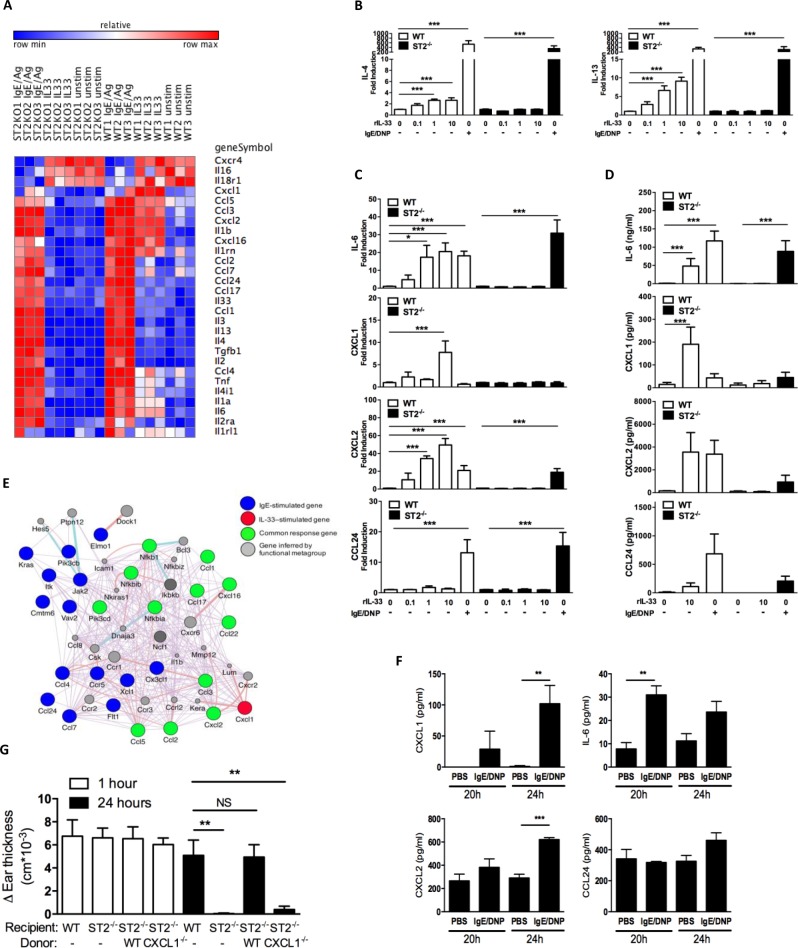
IgE-primed mast cells in peripheral tissues, including the skin, lung, and intestine, are key initiators of allergen-triggered edema and inflammation. Particularly in severe forms of allergy, this inflammation becomes strongly neutrophil dominated, and yet how mast cells coordinate this type of response is unknown. We and others have reported that activated mast cells--a hematopoietic cell type--can produce IL-33, a cytokine known to participate in allergic responses but generally considered as being of epithelial origin and driving Type 2 immune responses (e.g., ILC2 and eosinophil activation). Using models of skin anaphylaxis, our data reveal that mast cell-derived IL-33 also initiates neutrophilic inflammation. We demonstrate a cellular crosstalk mechanism whereby activated mast cells crosstalk to IL-33 receptor-bearing basophils, driving these basophils to adopt a unique response signature rich in neutrophil-associated molecules. We further establish that basophil expression of CXCL1 is necessary for IgE-driven neutrophilic inflammation. Our findings thus unearth a new mechanism by which mast cells initiate local inflammation after antigen triggering and might explain the complex inflammatory phenotypes observed in severe allergic diseases. Moreover, our findings (i) establish a functional link from IL-33 to neutrophilic inflammation that extends IL-33-mediated biology well beyond that of Type 2 immunity, and (ii) demonstrate the functional importance of hematopoietic cell-derived IL-33 in allergic pathogenesis.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: Paul J. Bryce is an employee of Sanofi-Genzyme and receives compensation from them. This study is independent of Sanofi-Genzyme and relates entirely to research conducted during his time at Northwestern University. This does not alter our adherence to PLOS ONE policies on sharing data and materials.
Figures

References
-
- Wershil BK, Wang ZS, Gordon JR, Galli SJ. Recruitment of neutrophils during IgE-dependent cutaneous late phase reactions in the mouse is mast cell-dependent. Partial inhibition of the reaction with antiserum against tumor necrosis factor-alpha. The Journal of clinical investigation. 1991;87(2):446–53. Epub 1991/02/01. 10.1172/JCI115016 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
