

Voltage Sensing in Bacterial Protein Translocation
- PMID: 31947864
- PMCID: PMC7023257
- DOI: 10.3390/biom10010078
Voltage Sensing in Bacterial Protein Translocation
Abstract
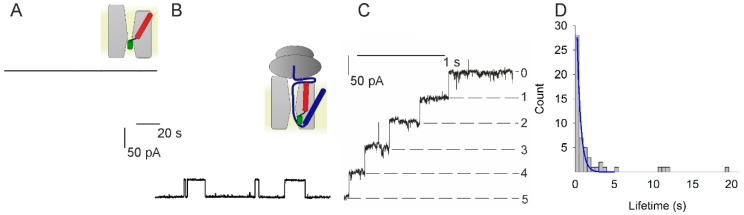
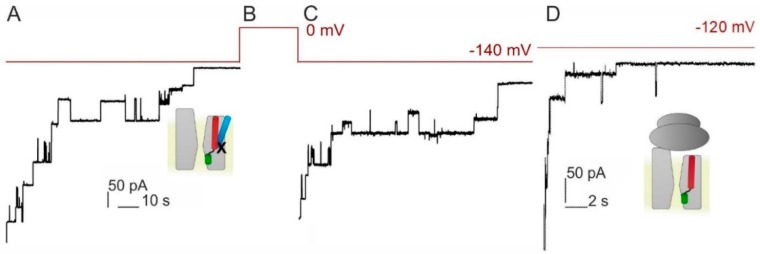
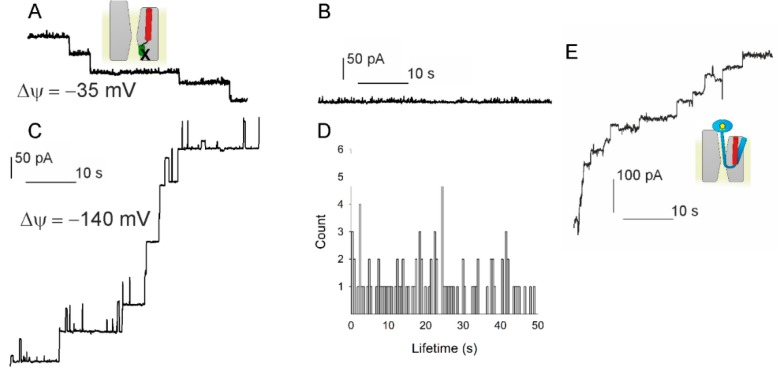
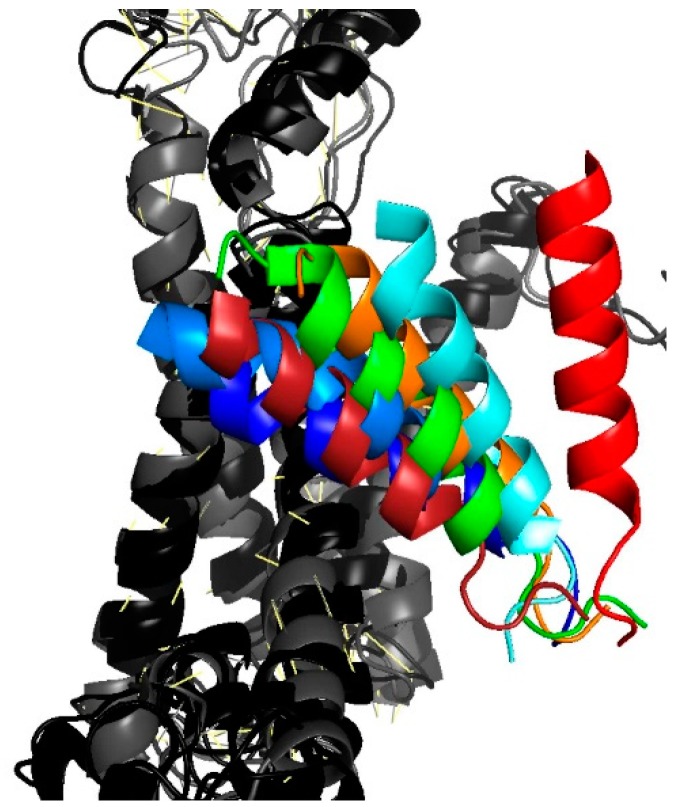
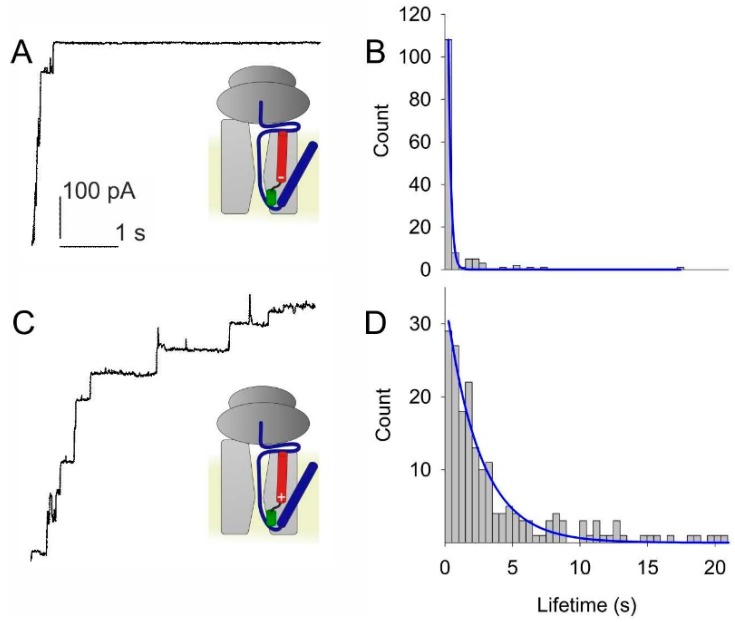
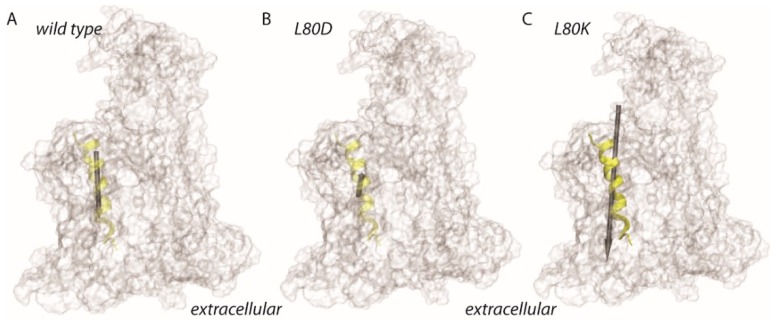
The bacterial channel SecYEG efficiently translocates both hydrophobic and hydrophilic proteins across the plasma membrane. Translocating polypeptide chains may dislodge the plug, a half helix that blocks the permeation of small molecules, from its position in the middle of the aqueous translocation channel. Instead of the plug, six isoleucines in the middle of the membrane supposedly seal the channel, by forming a gasket around the translocating polypeptide. However, this hypothesis does not explain how the tightness of the gasket may depend on membrane potential. Here, we demonstrate voltage-dependent closings of the purified and reconstituted channel in the presence of ligands, suggesting that voltage sensitivity may be conferred by motor protein SecA, ribosomes, signal peptides, and/or translocating peptides. Yet, the presence of a voltage sensor intrinsic to SecYEG was indicated by voltage driven closure of pores that were forced-open either by crosslinking the plug to SecE or by plug deletion. We tested the involvement of SecY's half-helix 2b (TM2b) in voltage sensing, since clearly identifiable gating charges are missing. The mutation L80D accelerated voltage driven closings by reversing TM2b's dipolar orientation. In contrast, the L80K mutation decelerated voltage induced closings by increasing TM2b's dipole moment. The observations suggest that TM2b is part of a larger voltage sensor. By partly aligning the combined dipole of this sensor with the orientation of the membrane-spanning electric field, voltage may drive channel closure.
Keywords: Sec61; SecY; gating; translocon.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Seinen A.B., Driessen A.J.M. Single-molecule studies on the protein translocon. In: Dill K.A., editor. Annual Review of Biophysics, 48. Annual Reviews; Palo Alto, CA, USA: 2019. pp. 185–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
