

N 6-methyladenosine of chromosome-associated regulatory RNA regulates chromatin state and transcription
- PMID: 31949099
- PMCID: PMC7213019
- DOI: 10.1126/science.aay6018
N 6-methyladenosine of chromosome-associated regulatory RNA regulates chromatin state and transcription
Abstract
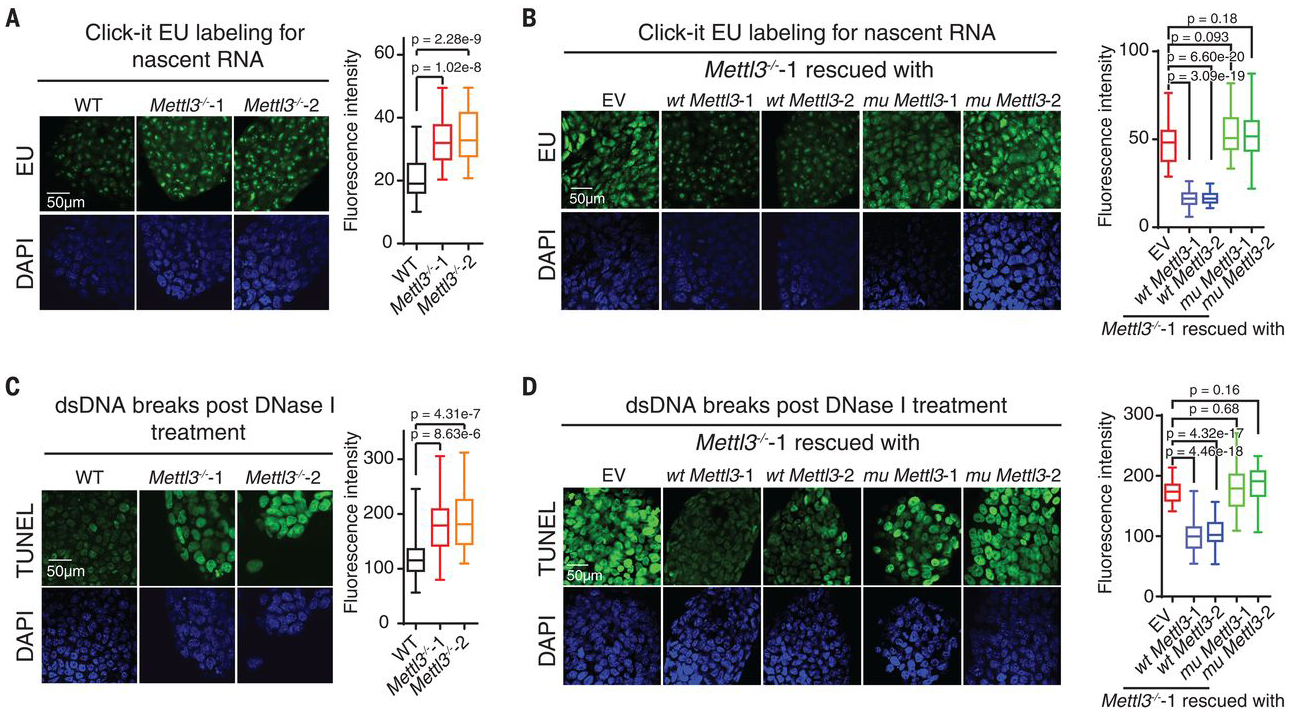
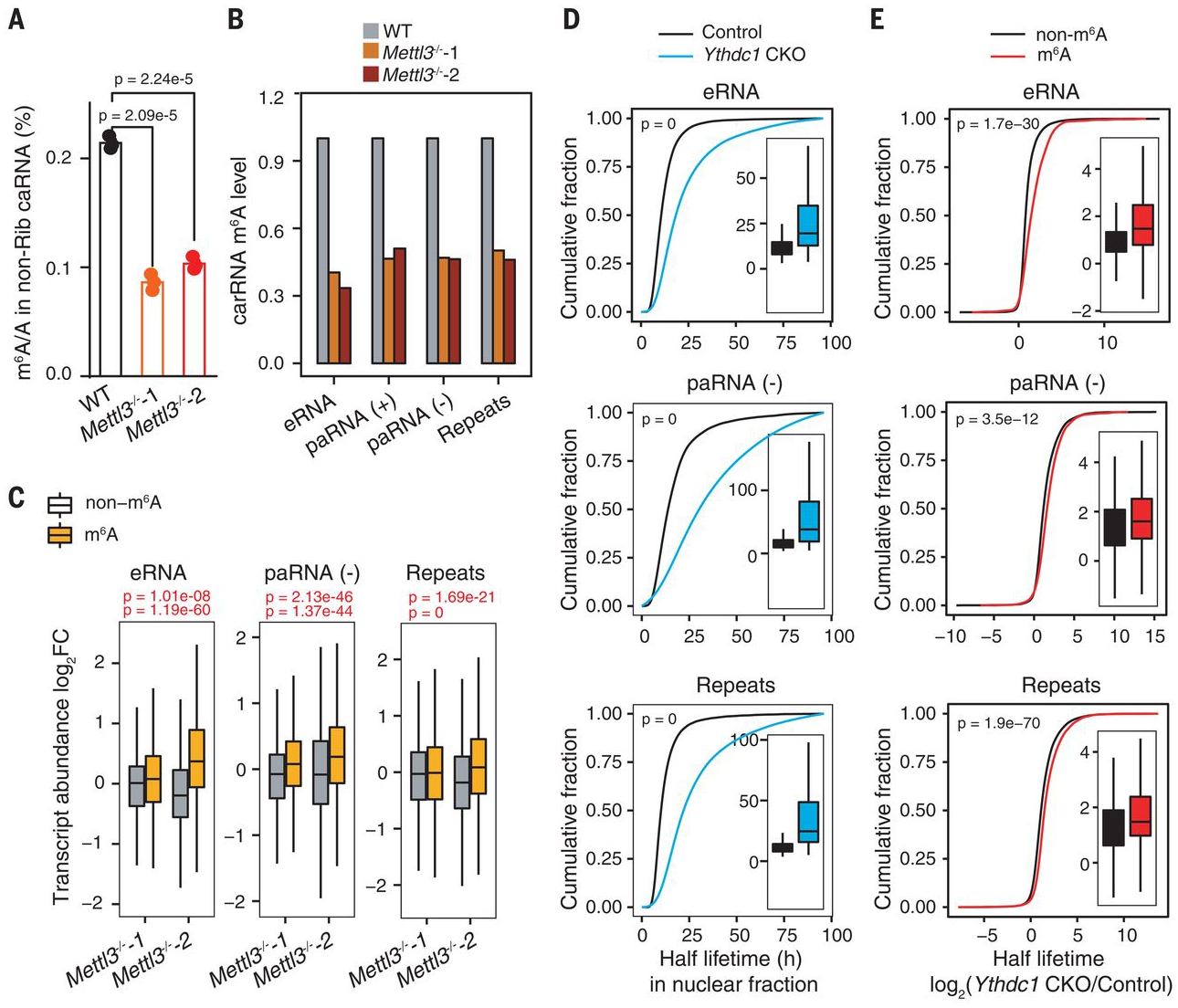
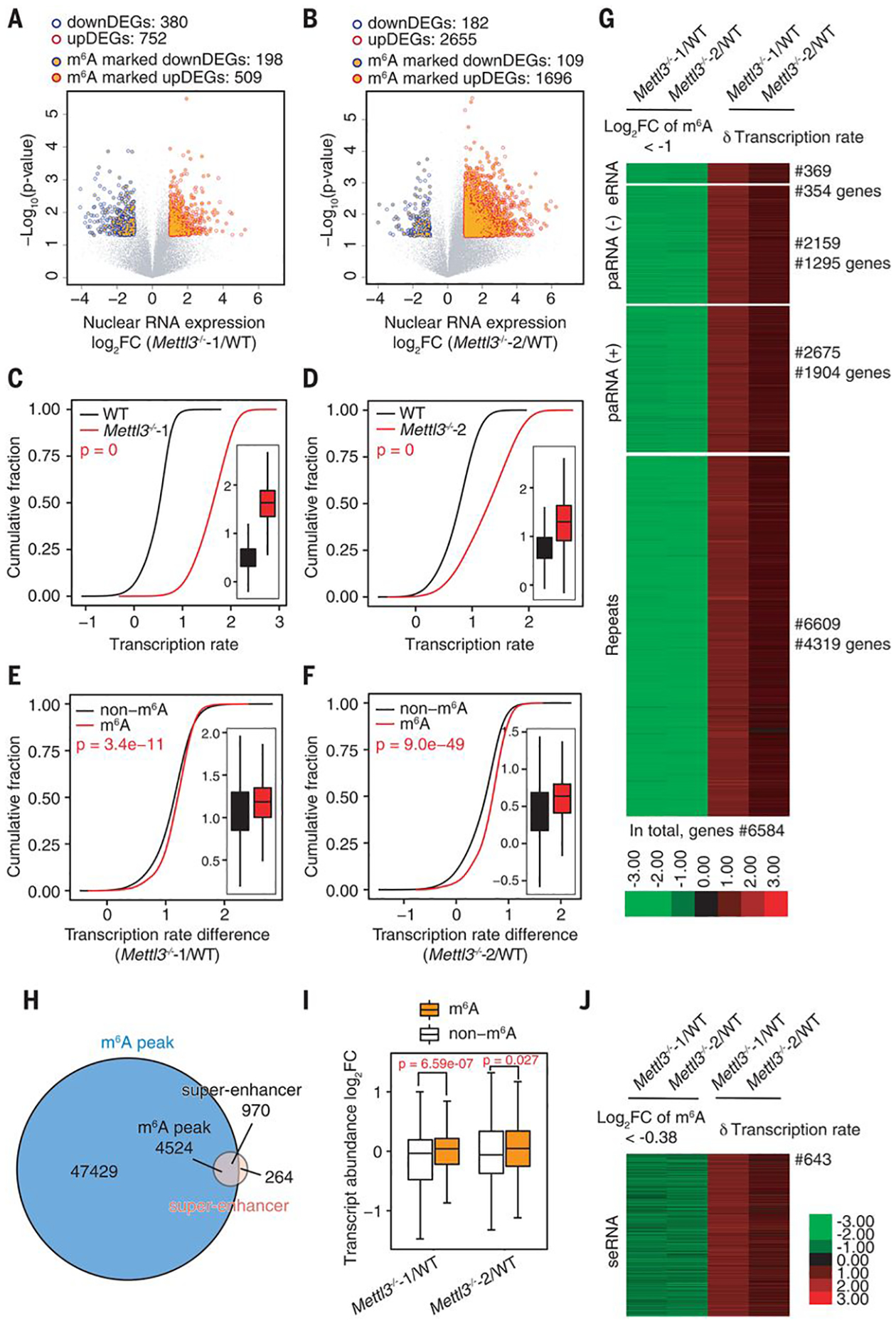
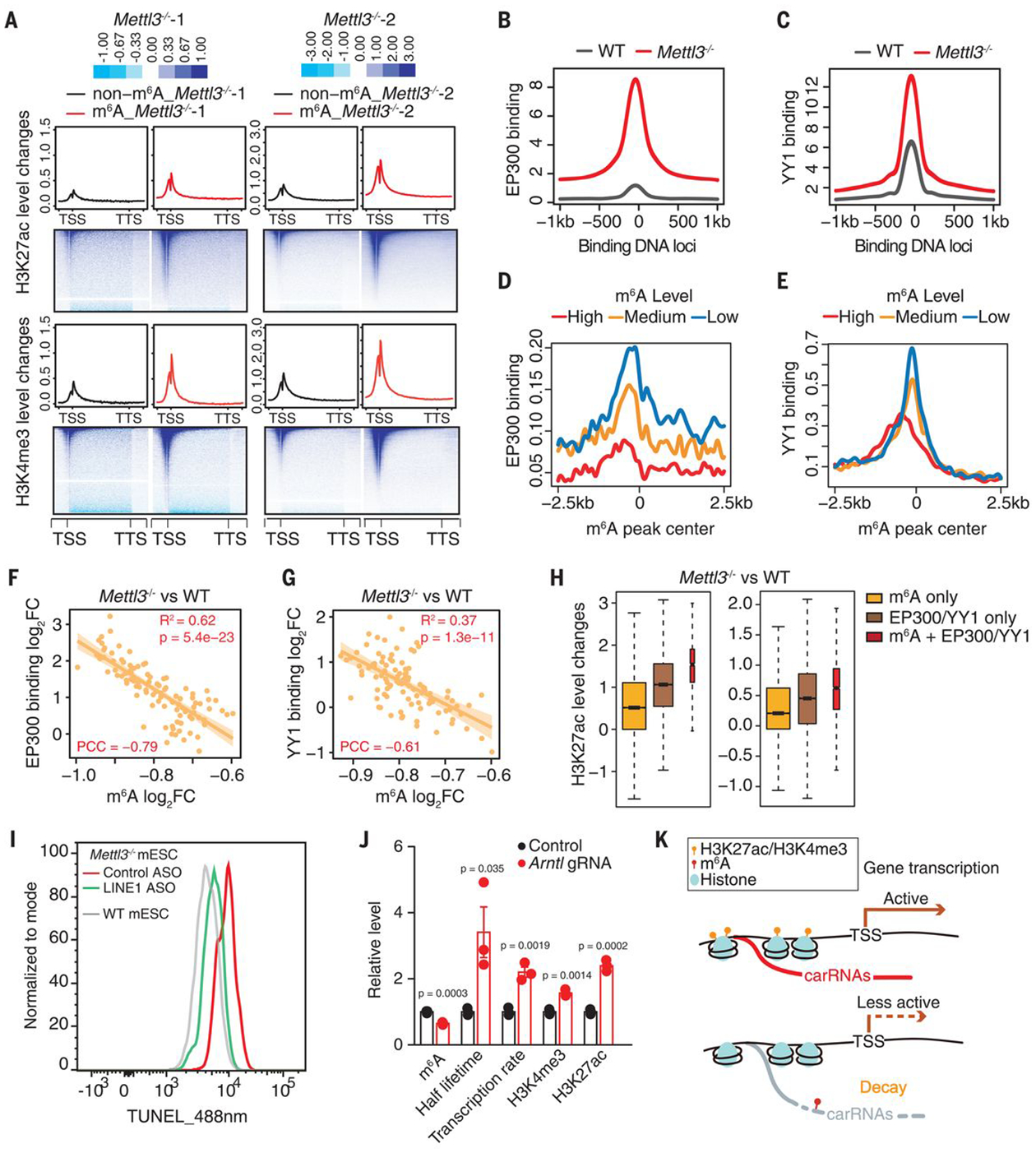
N 6-methyladenosine (m6A) regulates stability and translation of messenger RNA (mRNA) in various biological processes. In this work, we show that knockout of the m6A writer Mettl3 or the nuclear reader Ythdc1 in mouse embryonic stem cells increases chromatin accessibility and activates transcription in an m6A-dependent manner. We found that METTL3 deposits m6A modifications on chromosome-associated regulatory RNAs (carRNAs), including promoter-associated RNAs, enhancer RNAs, and repeat RNAs. YTHDC1 facilitates the decay of a subset of these m6A-modified RNAs, especially elements of the long interspersed element-1 family, through the nuclear exosome targeting-mediated nuclear degradation. Reducing m6A methylation by METTL3 depletion or site-specific m6A demethylation of selected carRNAs elevates the levels of carRNAs and promotes open chromatin state and downstream transcription. Collectively, our results reveal that m6A on carRNAs can globally tune chromatin state and transcription.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Dominissini D et al., Nature 485, 201–206 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
