

Fyn tyrosine kinase, a downstream target of receptor tyrosine kinases, modulates antiglioma immune responses
- PMID: 31950181
- PMCID: PMC7283034
- DOI: 10.1093/neuonc/noaa006
Fyn tyrosine kinase, a downstream target of receptor tyrosine kinases, modulates antiglioma immune responses
Erratum in
-
Erratum: Fyn tyrosine kinase, a downstream target of receptor tyrosine kinases, modulates antiglioma immune responses.Neuro Oncol. 2021 Aug 2;23(8):1413. doi: 10.1093/neuonc/noaa063. Neuro Oncol. 2021. PMID: 32428217 Free PMC article. No abstract available.
Abstract
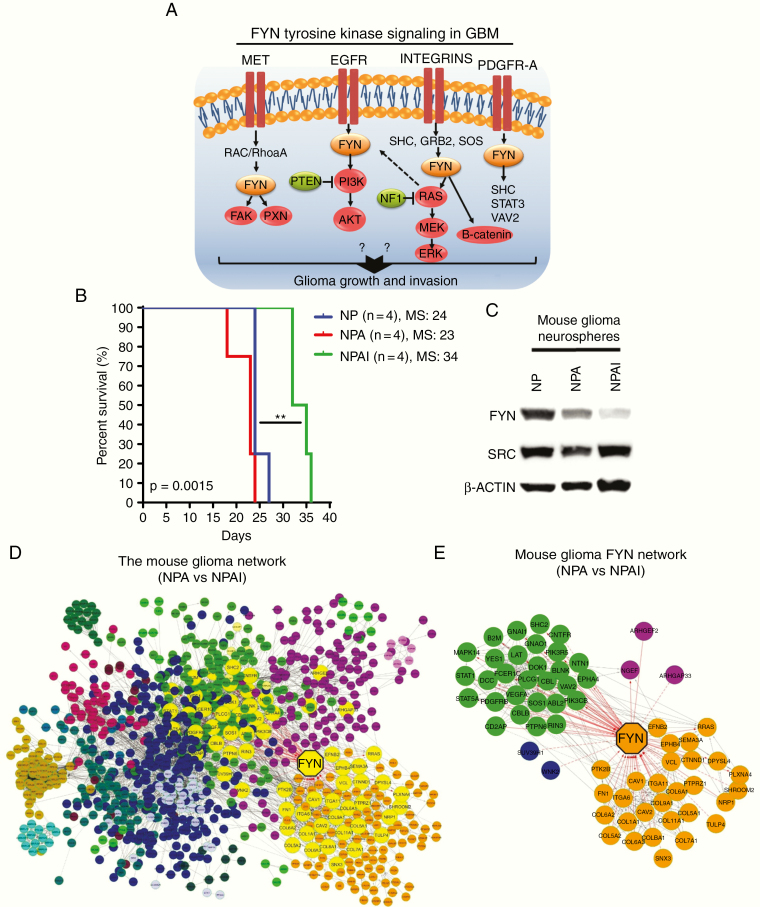
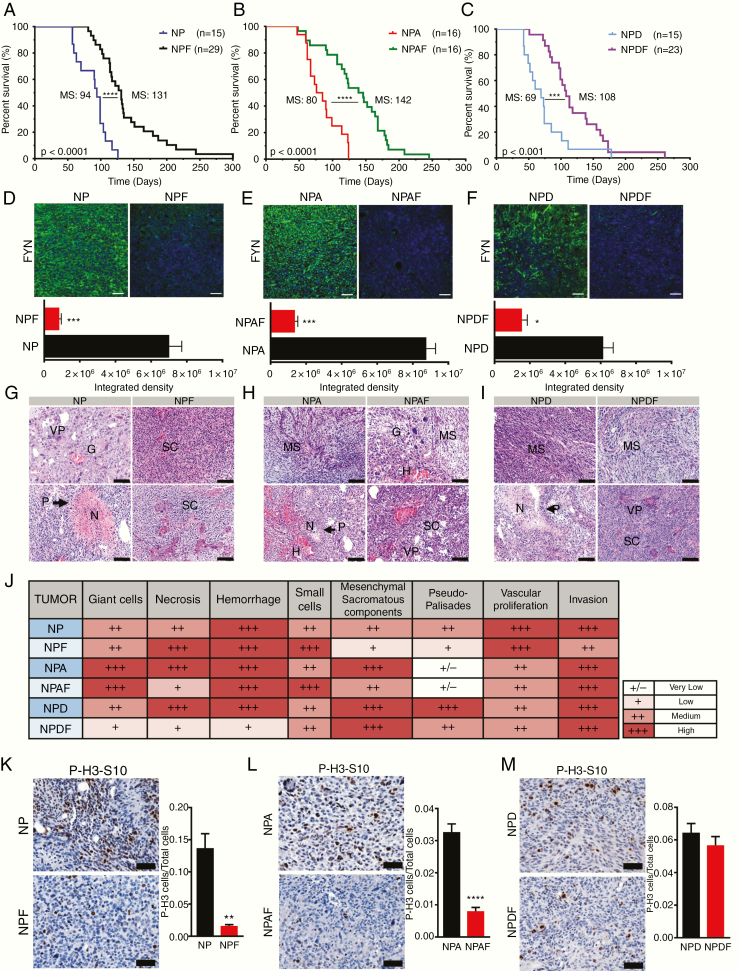
Background: High-grade gliomas are aggressive and immunosuppressive brain tumors. Molecular mechanisms that regulate the inhibitory immune tumor microenvironment (TME) and glioma progression remain poorly understood. Fyn tyrosine kinase is a downstream target of the oncogenic receptor tyrosine kinase pathway and is overexpressed in human gliomas. Fyn's role in vivo in glioma growth remains unknown. We investigated whether Fyn regulates glioma initiation, growth and invasion.
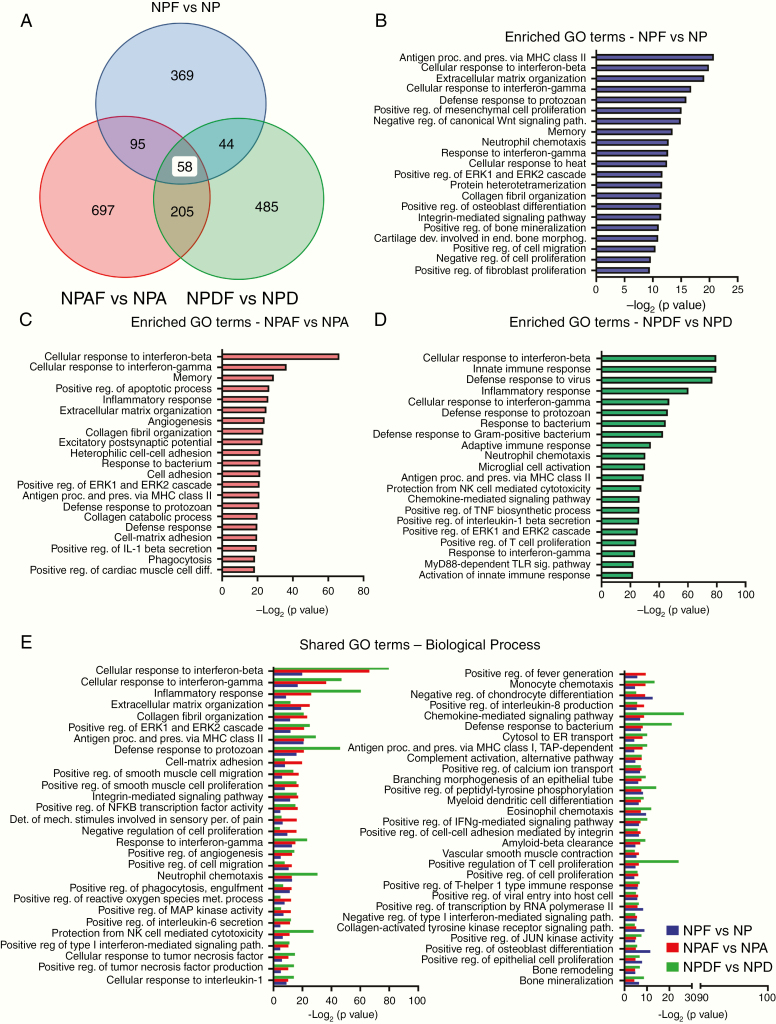
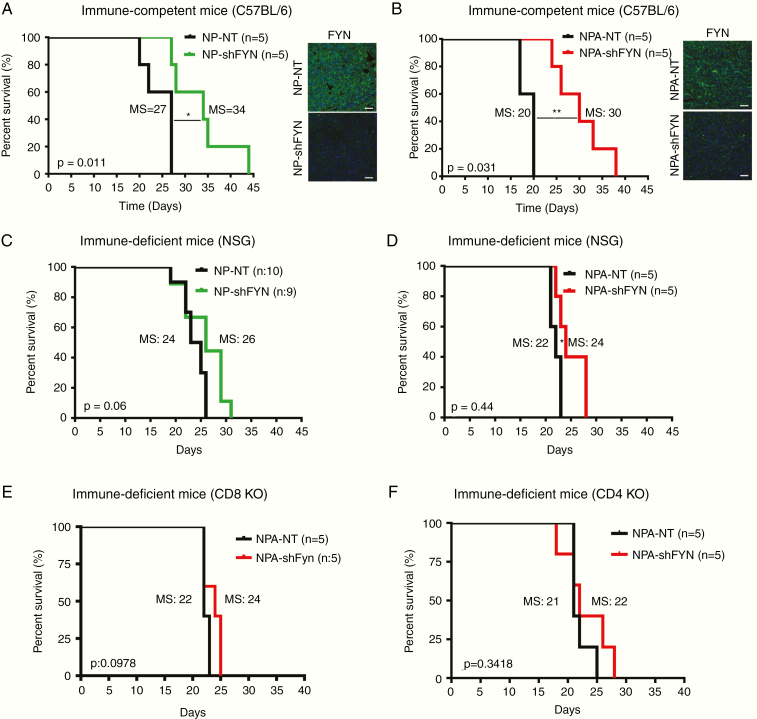
Methods: We evaluated the role of Fyn using genetically engineered mouse glioma models (GEMMs). We also generated Fyn knockdown stem cells to induce gliomas in immune-competent and immune-deficient mice (nonobese diabetic severe combined immunodeficient gamma mice [NSG], CD8-/-, CD4-/-). We analyzed molecular mechanism by RNA sequencing and bioinformatics analysis. Flow cytometry was used to characterize immune cellular infiltrates in the Fyn knockdown glioma TME.
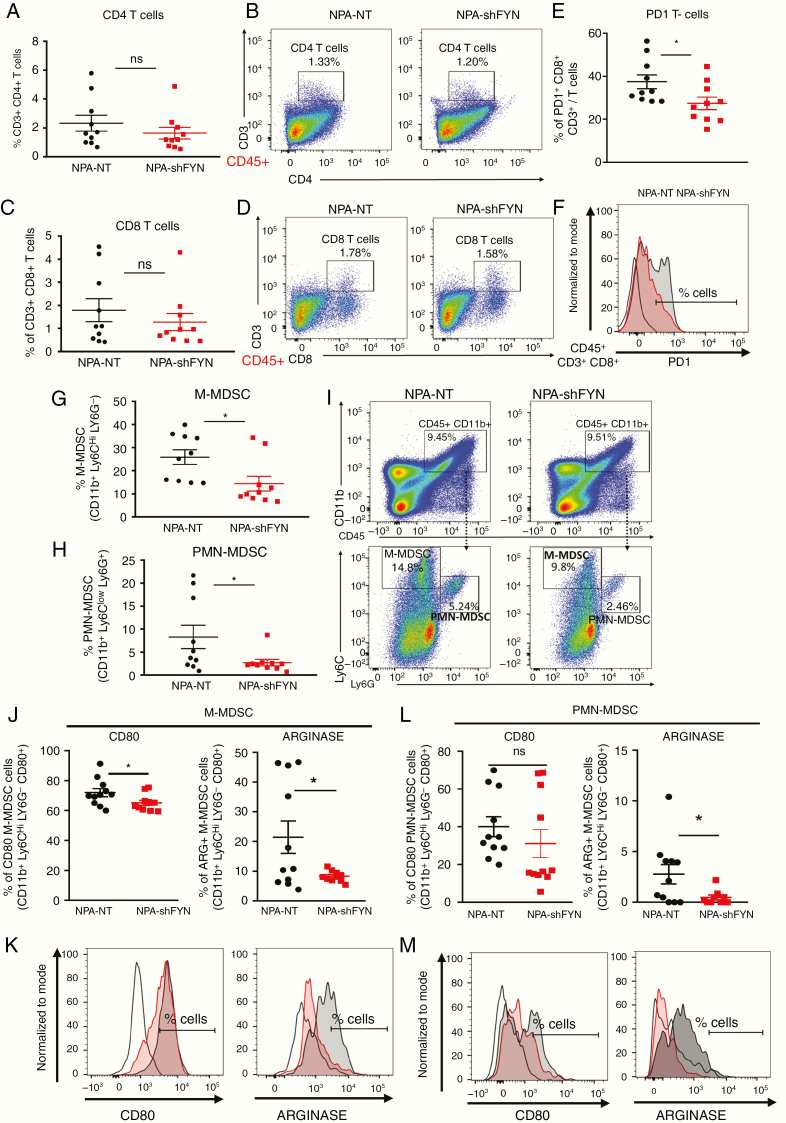
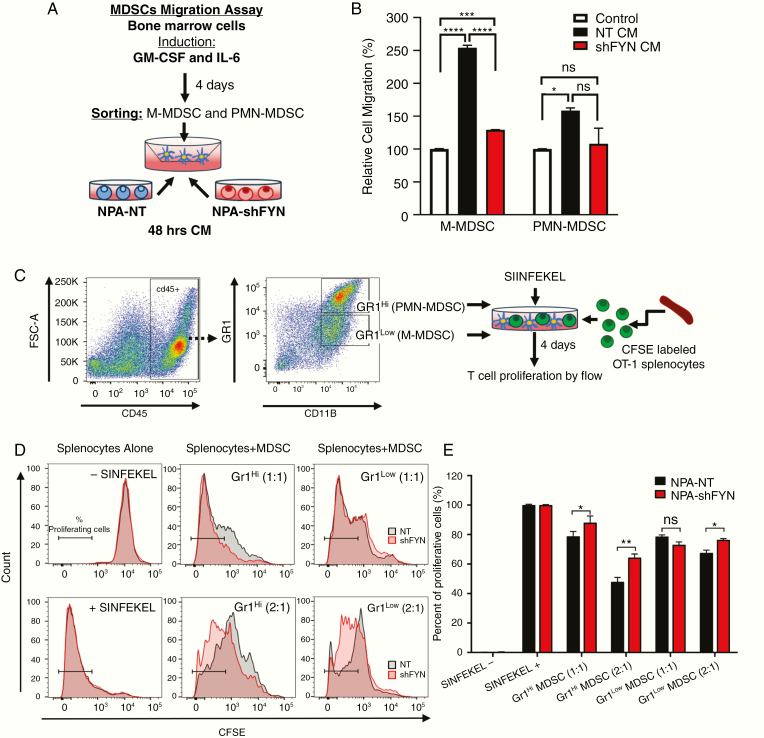
Results: We demonstrate that Fyn knockdown in diverse immune-competent GEMMs of glioma reduced tumor progression and significantly increased survival. Gene ontology (GO) analysis of differentially expressed genes in wild-type versus Fyn knockdown gliomas showed enrichment of GOs related to immune reactivity. However, in NSG and CD8-/- and CD4-/- immune-deficient mice, Fyn knockdown gliomas failed to show differences in survival. These data suggest that the expression of Fyn in glioma cells reduces antiglioma immune activation. Examination of glioma immune infiltrates by flow cytometry displayed reduction in the amount and activity of immune suppressive myeloid derived cells in the Fyn glioma TME.
Conclusions: Gliomas employ Fyn mediated mechanisms to enhance immune suppression and promote tumor progression. We propose that Fyn inhibition within glioma cells could improve the efficacy of antiglioma immunotherapies.
Keywords: Fyn tyrosine kinase; antitumor immune responses; glioma; myeloid-derived suppressor cells.
© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Neuro-Oncology. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
Comment in
-
A Fyn romance: tumor cell Fyn kinase suppresses the immune microenvironment.Neuro Oncol. 2020 Jun 9;22(6):746-747. doi: 10.1093/neuonc/noaa082. Neuro Oncol. 2020. PMID: 32227231 Free PMC article. No abstract available.
References
-
- Reifenberger G, Wirsching HG, Knobbe-Thomsen CB, Weller M. Advances in the molecular genetics of gliomas—implications for classification and therapy. Nat Rev Clin Oncol. 2017;14(7):434–452. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
