

An RNA Thermometer Activity of the West Nile Virus Genomic 3'-Terminal Stem-Loop Element Modulates Viral Replication Efficiency during Host Switching
- PMID: 31952291
- PMCID: PMC7019923
- DOI: 10.3390/v12010104
An RNA Thermometer Activity of the West Nile Virus Genomic 3'-Terminal Stem-Loop Element Modulates Viral Replication Efficiency during Host Switching
Abstract
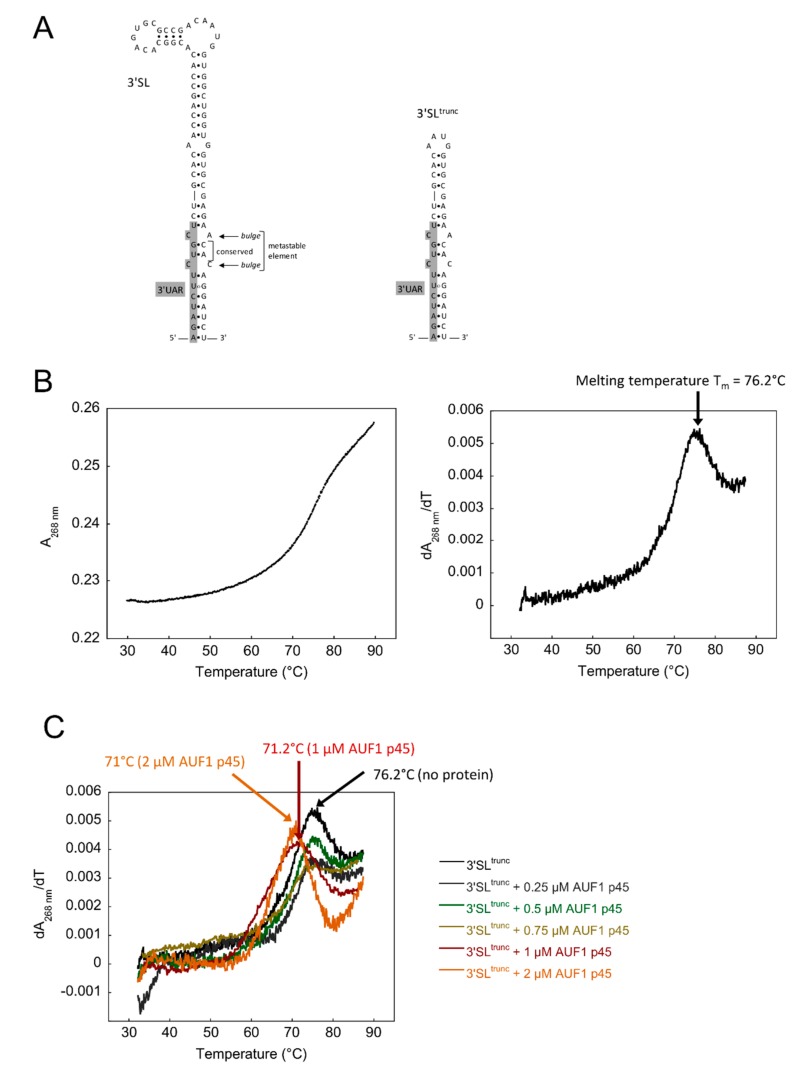
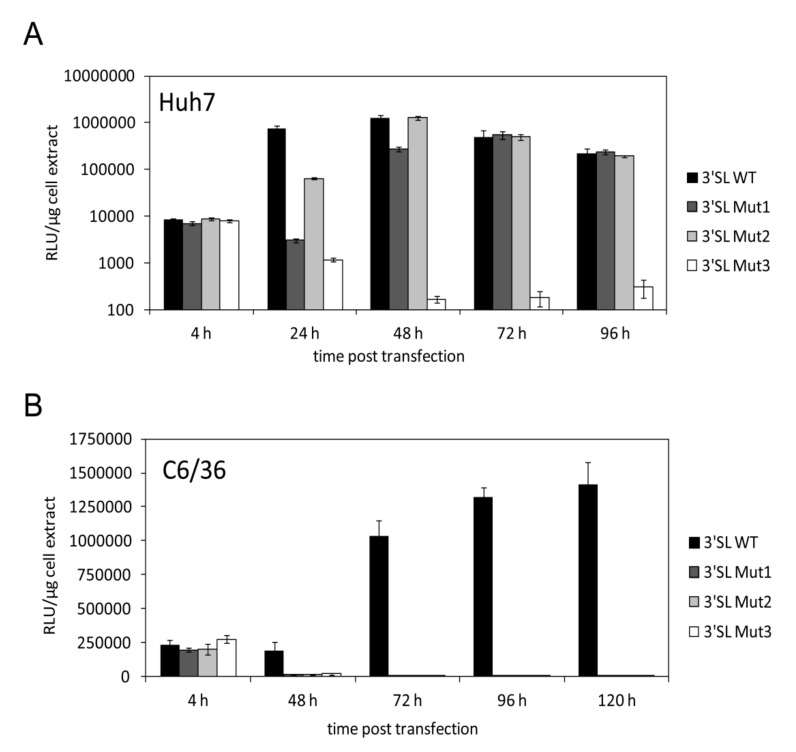
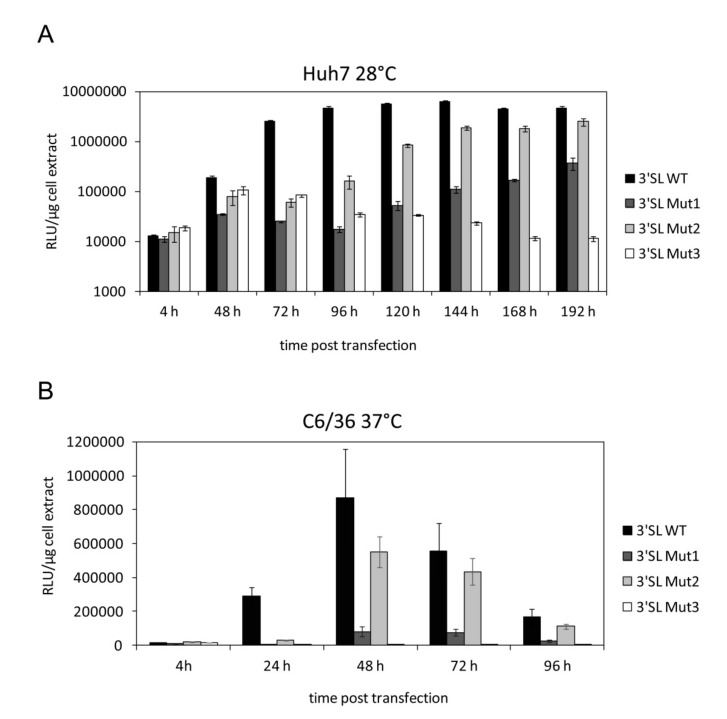
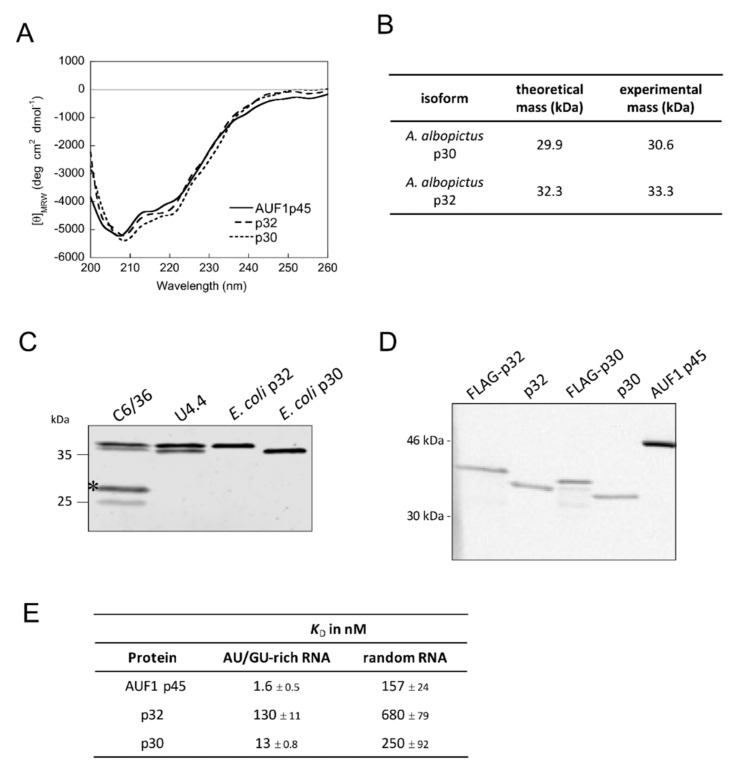
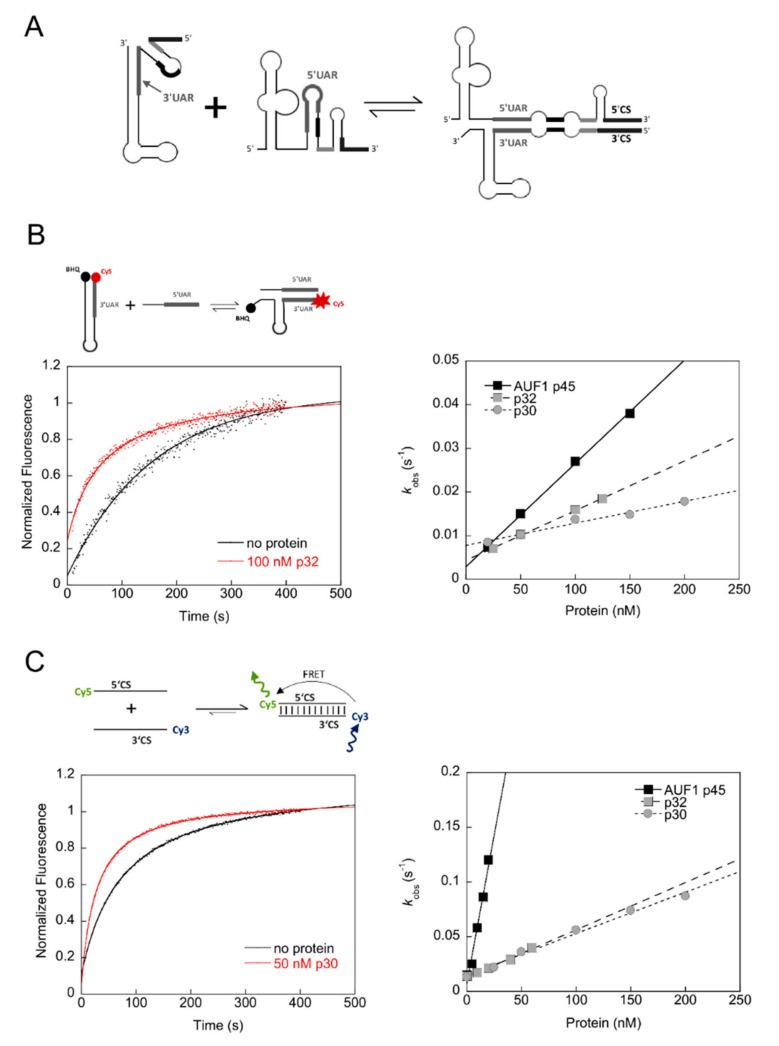
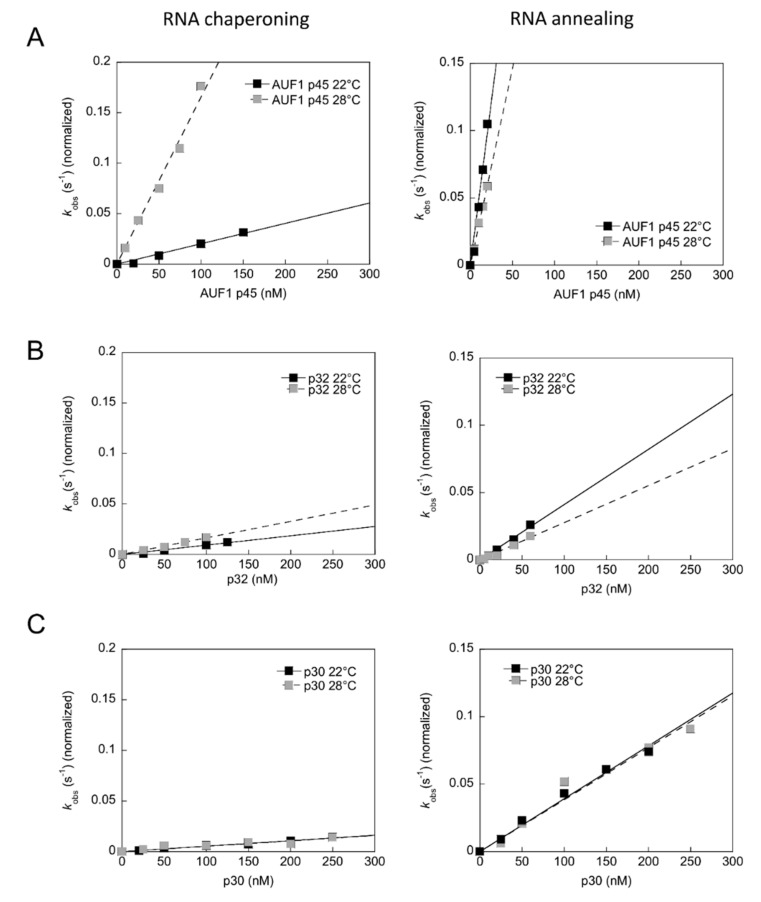
The 3'-terminal stem-loop (3'SL) of the RNA genome of the flavivirus West Nile (WNV) harbors, in its stem, one of the sequence elements that are required for genome cyclization. As cyclization is a prerequisite for the initiation of viral replication, the 3'SL was proposed to act as a replication silencer. The lower part of the 3'SL is metastable and confers a structural flexibility that may regulate the switch from the linear to the circular conformation of the viral RNA. In the human system, we previously demonstrated that a cellular RNA-binding protein, AUF1 p45, destabilizes the 3'SL, exposes the cyclization sequence, and thus promotes flaviviral genome cyclization and RNA replication. By investigating mutant RNAs with increased 3'SL stabilities, we showed the specific conformation of the metastable element to be a critical determinant of the helix-destabilizing RNA chaperone activity of AUF1 p45 and of the precision and efficiency of the AUF1 p45-supported initiation of RNA replication. Studies of stability-increasing mutant WNV replicons in human and mosquito cells revealed that the cultivation temperature considerably affected the replication efficiencies of the viral RNA variants and demonstrated the silencing effect of the 3'SL to be temperature dependent. Furthermore, we identified and characterized mosquito proteins displaying similar activities as AUF1 p45. However, as the RNA remodeling activities of the mosquito proteins were found to be considerably lower than those of the human protein, a potential cell protein-mediated destabilization of the 3'SL was suggested to be less efficient in mosquito cells. In summary, our data support a model in which the 3'SL acts as an RNA thermometer that modulates flavivirus replication during host switching.
Keywords: Flavivirus; RNA annealer; RNA chaperone; RNA remodeling; RNA replication; RNA structure; RNA thermometer; West Nile virus; host factor; host switching.
Conflict of interest statement
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
