

Astrocyte-T cell crosstalk regulates region-specific neuroinflammation
- PMID: 31961459
- PMCID: PMC7317491
- DOI: 10.1002/glia.23783
Astrocyte-T cell crosstalk regulates region-specific neuroinflammation
Abstract
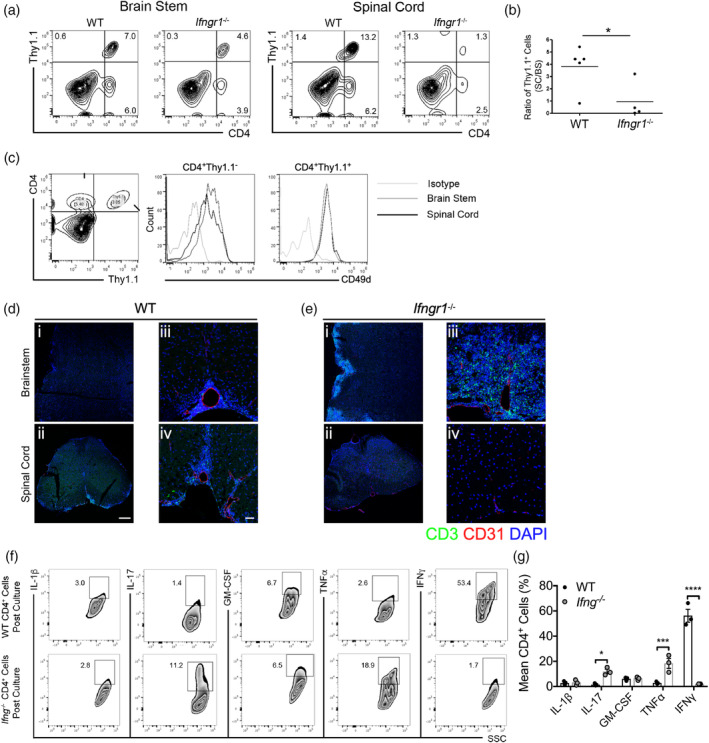
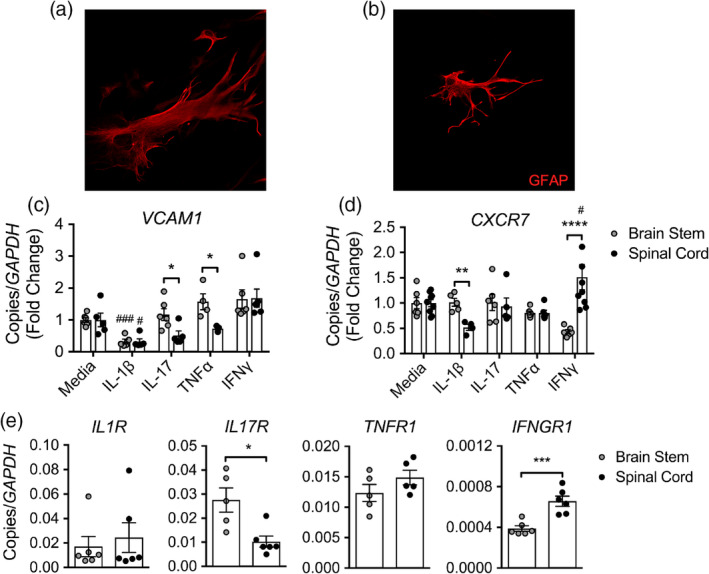
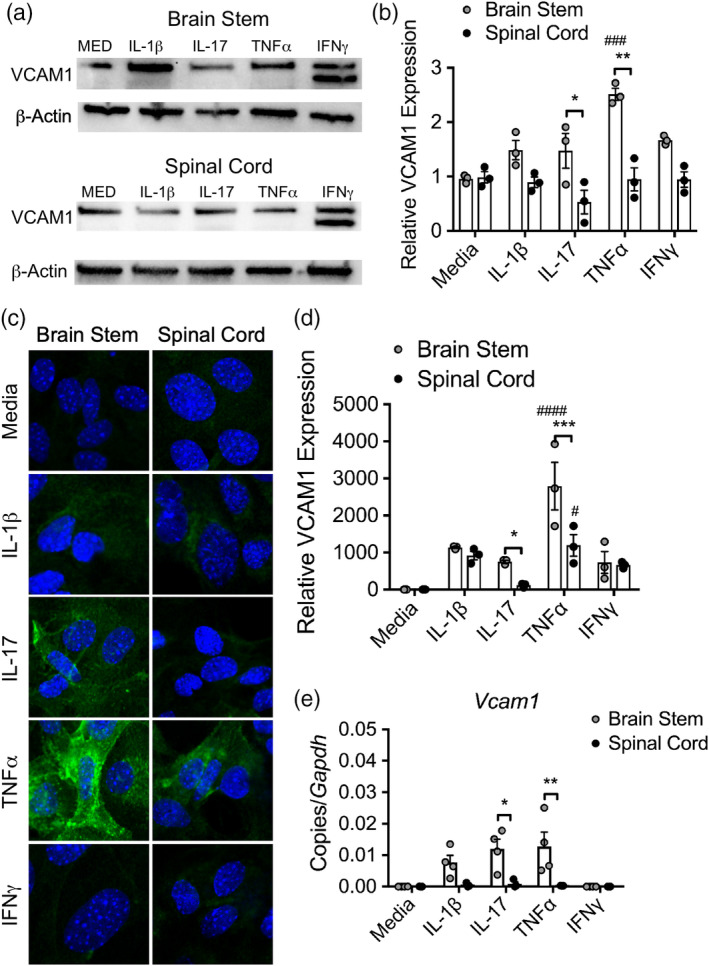
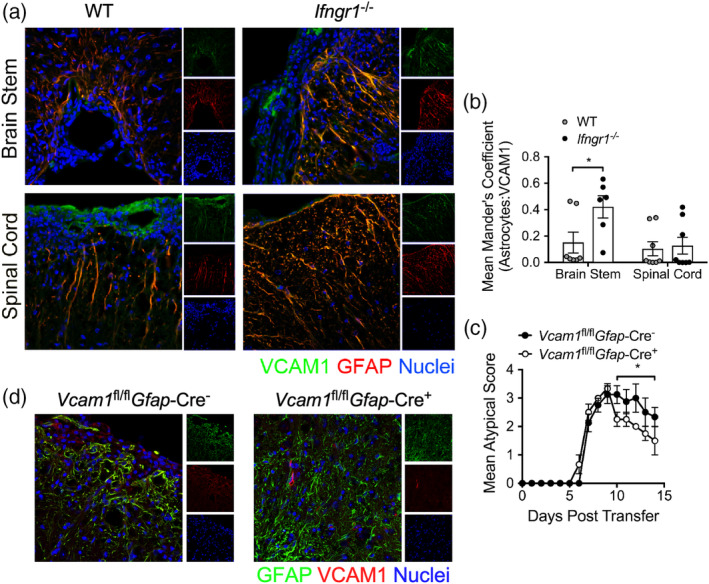
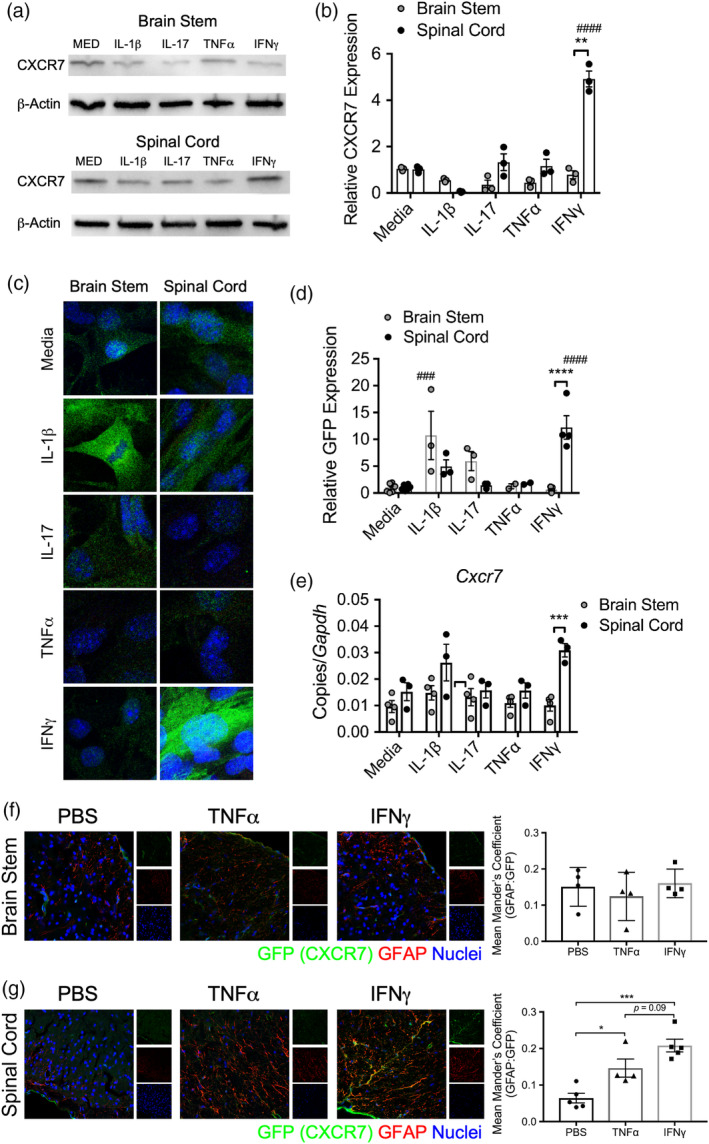
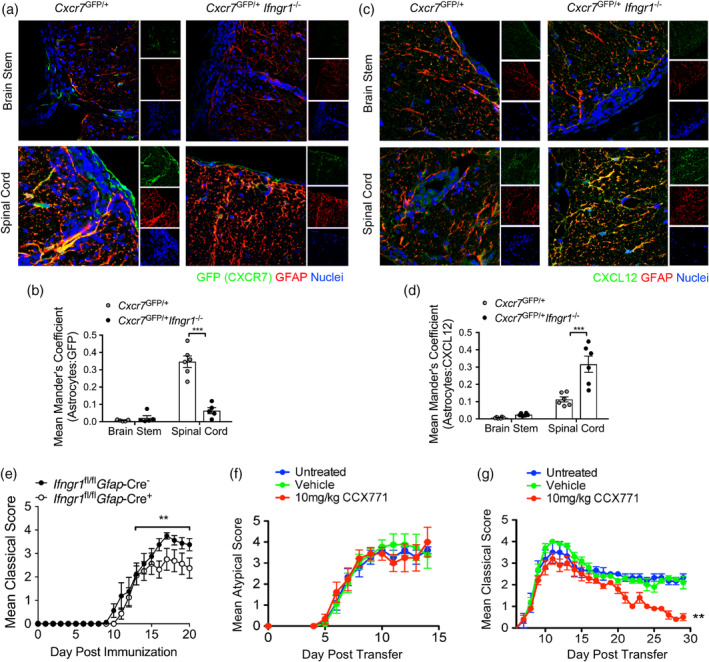
During multiple sclerosis (MS), an inflammatory and neurodegenerative disease of the central nervous system (CNS), symptoms, and outcomes are determined by the location of inflammatory lesions. While we and others have shown that T cell cytokines differentially regulate leukocyte entry into perivascular spaces and regional parenchymal localization in murine models of MS, the molecular mechanisms of this latter process are poorly understood. Here, we demonstrate that astrocytes exhibit region-specific responses to T cell cytokines that promote hindbrain versus spinal cord neuroinflammation. Analysis of cytokine receptor expression in human astrocytes showed region-specific responsiveness to Th1 and Th17 inflammatory cytokines. Consistent with this, human and murine astrocytes treated with these cytokines exhibit differential expression of the T cell localizing molecules VCAM-1 and CXCR7 that is both cytokine and CNS region-specific. Using in vivo models of spinal cord versus brain stem trafficking of myelin-specific T cells and astrocyte-specific deletion strategies, we confirmed that Th1 and Th17 cytokines differentially regulate astrocyte expression of VCAM-1 and CXCR7 in these locations. Finally, stereotaxic injection of individual cytokines into the hindbrain or spinal cord revealed region- and cytokine-specific modulation of localizing cue expression by astrocytes. These findings identify a role for inflammatory cytokines in mediating local astrocyte-dependent mechanisms of immune cell trafficking within the CNS during neuroinflammation.
Keywords: CXCR7; T cell; VCAM-1; astrocyte; cytokine; neuroinflammation; regional heterogeneity.
© 2020 The Authors. Glia published by Wiley Periodicals, Inc.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Bever, C. T., Jr. , Panitch, H. S. , Levy, H. B. , McFarlin, D. E. , & Johnson, K. P. (1991). Gamma‐interferon induction in patients with chronic progressive MS. Neurology, 41(7), 1124–1127. - PubMed
-
- Billiau, A. , Heremans, H. , Vandekerckhove, F. , Dijkmans, R. , Sobis, H. , Meulepas, E. , & Carton, H. (1988). Enhancement of experimental allergic encephalomyelitis in mice by antibodies against IFN‐gamma. Journal of Immunology, 140(5), 1506–1510. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
