

Analysis of variants in GATA4 and FOG2/ZFPM2 demonstrates benign contribution to 46,XY disorders of sex development
- PMID: 31962012
- PMCID: PMC7057099
- DOI: 10.1002/mgg3.1095
Analysis of variants in GATA4 and FOG2/ZFPM2 demonstrates benign contribution to 46,XY disorders of sex development
Abstract
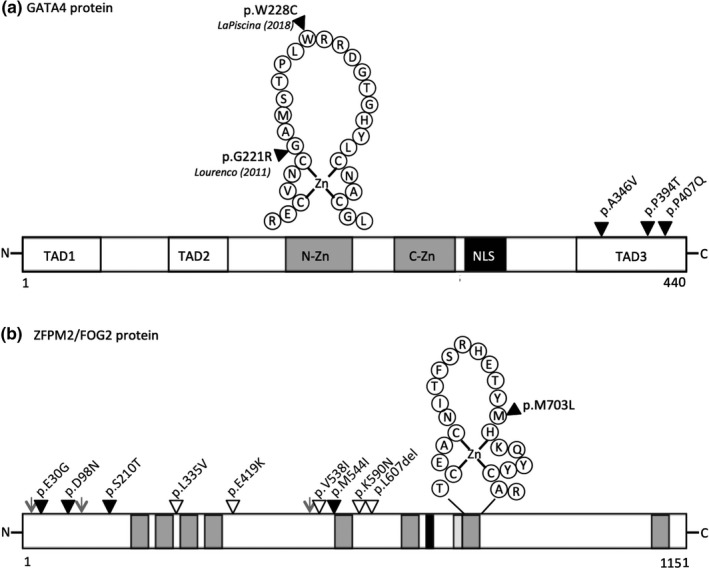
Background: GATA-binding protein 4 (GATA4) and Friend of GATA 2 protein (FOG2, also known as ZFPM2) form a heterodimer complex that has been shown to influence transcription of genes in a number of developmental systems. Recent evidence has also shown these genes play a role in gonadal sexual differentiation in humans. Previously we identified four variants in GATA4 and an unexpectedly large number of variants in ZFPM2 in a cohort of individuals with 46,XY Differences/Disorders of Sex Development (DSD) (Eggers et al, Genome Biology, 2016; 17: 243).
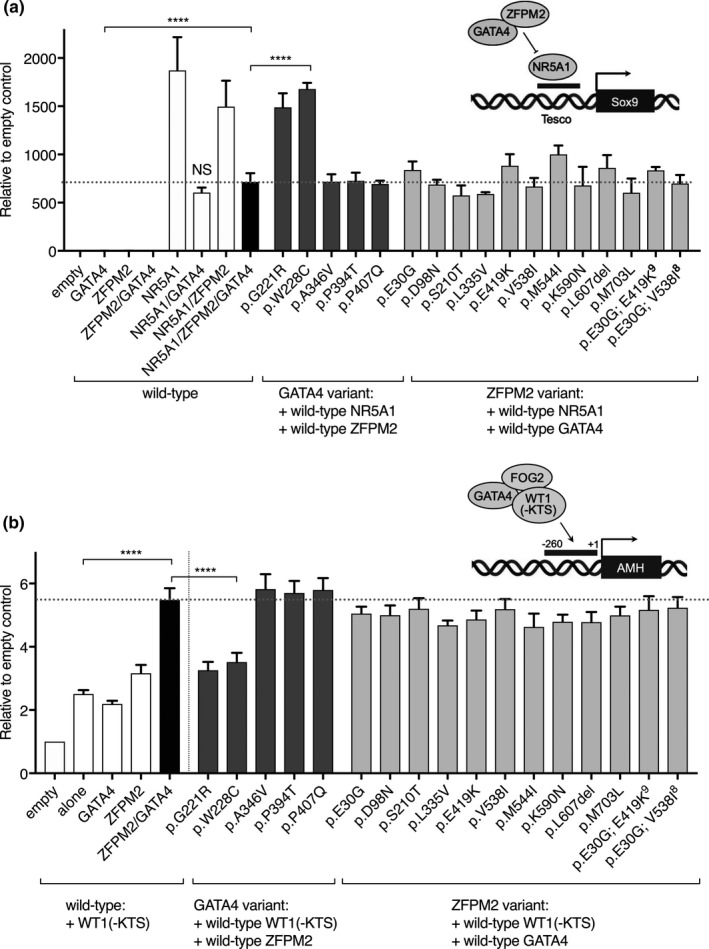
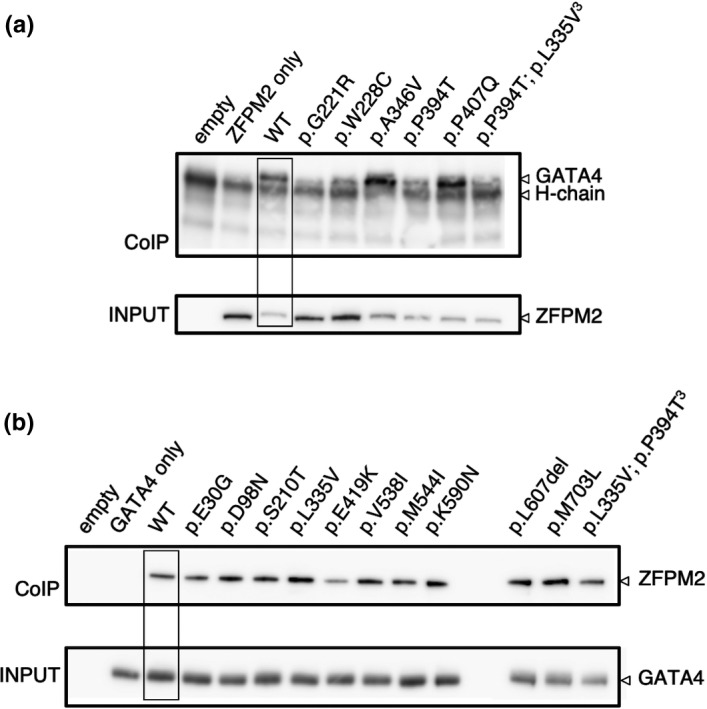
Method: Here, we review variant curation and test the functional activity of GATA4 and ZFPM2 variants. We assess variant transcriptional activity on gonadal specific promoters (Sox9 and AMH) and variant protein-protein interactions.
Results: Our findings support that the majority of GATA4 and ZFPM2 variants we identified are benign in their contribution to 46,XY DSD. Indeed, only one variant, in the conserved N-terminal zinc finger of GATA4, was considered pathogenic, with functional analysis confirming differences in its ability to regulate Sox9 and AMH and in protein interaction with ZFPM2.
Conclusions: Our study helps define the genetic factors contributing to 46,XY DSD and suggests that the majority of variants we identified in GATA4 and ZFPM2/FOG2 are not causative.
Keywords: FOG2; GATA4; ZFPM2; disorders of sexual development; functional analysis; mutations.
© 2019 The Murdoch Children's Research Institute. Molecular Genetics & Genomic Medicine published by Wiley Periodicals, Inc.
Conflict of interest statement
Dr Davis reports having received honoraria from Besins Healthcare and Pfizer Australia and has been a consultant to Mayne Pharmaceuticals, Lawley Pharmaceuticals and Que Oncology. All other authors declare no conflict of interest.
Figures
Similar articles
-
Variants of STAR, AMH and ZFPM2/FOG2 May Contribute towards the Broad Phenotype Observed in 46,XY DSD Patients with Heterozygous Variants of NR5A1.Int J Mol Sci. 2020 Nov 13;21(22):8554. doi: 10.3390/ijms21228554. Int J Mol Sci. 2020. PMID: 33202802 Free PMC article.
-
Correct dosage of Fog2 and Gata4 transcription factors is critical for fetal testis development in mice.Proc Natl Acad Sci U S A. 2007 Sep 18;104(38):14994-9. doi: 10.1073/pnas.0701677104. Epub 2007 Sep 11. Proc Natl Acad Sci U S A. 2007. PMID: 17848526 Free PMC article.
-
Mutations in the FOG2/ZFPM2 gene are associated with anomalies of human testis determination.Hum Mol Genet. 2014 Jul 15;23(14):3657-65. doi: 10.1093/hmg/ddu074. Epub 2014 Feb 18. Hum Mol Genet. 2014. PMID: 24549039
-
Oligogenic Causes of Human Differences of Sex Development: Facing the Challenge of Genetic Complexity.Horm Res Paediatr. 2023;96(2):169-179. doi: 10.1159/000519691. Epub 2021 Sep 17. Horm Res Paediatr. 2023. PMID: 34537773 Review.
-
GATA-4 Variants in Two Unrelated Cases with 46, XY Disorder of Sex Development and Review of the Literature.J Clin Res Pediatr Endocrinol. 2022 Dec 1;14(4):469-474. doi: 10.4274/jcrpe.galenos.2021.2021.0112. Epub 2021 Aug 6. J Clin Res Pediatr Endocrinol. 2022. PMID: 34355877 Free PMC article. Review.
Cited by
-
Insights Into the Roles of GATA Factors in Mammalian Testis Development and the Control of Fetal Testis Gene Expression.Front Endocrinol (Lausanne). 2022 May 26;13:902198. doi: 10.3389/fendo.2022.902198. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 35692407 Free PMC article. Review.
-
Whole-genome sequencing identifies new candidate genes for nonobstructive azoospermia.Andrology. 2022 Nov;10(8):1605-1624. doi: 10.1111/andr.13269. Epub 2022 Sep 7. Andrology. 2022. PMID: 36017582 Free PMC article.
-
Variants of STAR, AMH and ZFPM2/FOG2 May Contribute towards the Broad Phenotype Observed in 46,XY DSD Patients with Heterozygous Variants of NR5A1.Int J Mol Sci. 2020 Nov 13;21(22):8554. doi: 10.3390/ijms21228554. Int J Mol Sci. 2020. PMID: 33202802 Free PMC article.
-
A Rare Case of Precocious Puberty in a Child with a Novel GATA-4 Gene Mutation: Implications for Disorders of Sex Development (DSD) and Review of the Literature.Genes (Basel). 2023 Aug 16;14(8):1631. doi: 10.3390/genes14081631. Genes (Basel). 2023. PMID: 37628683 Free PMC article. Review.
-
Molecular and Cytogenetic Analysis of Romanian Patients with Differences in Sex Development.Diagnostics (Basel). 2021 Nov 14;11(11):2107. doi: 10.3390/diagnostics11112107. Diagnostics (Basel). 2021. PMID: 34829455 Free PMC article.
References
-
- Abilash, V. G. , Radha, S. , Marimuthu, K. M. , Thangaraj, K. , Arun, S. , Nishu, S. , … Anuradha, D. (2016). Clinical, cytogenetic and molecular analysis of androgen insensitivity syndromes from south Indian cohort and detection and in‐silico characterization of androgen receptor gene mutations. Clinica Chimica Acta, 453, 123–130. 10.1016/j.cca.2015.12.012 - DOI - PubMed
-
- Ahmed, S. F. , Dobbie, R. , Finlayson, A. R. , Gilbert, J. , Youngson, G. , Chalmers, J. , & Stone, D. (2004). Prevalence of hypospadias and other genital anomalies among singleton births, 1988–1997, in Scotland. Archives of Disease in Childhood ‐ Fetal and Neonatal Edition, 89, F149–F151. 10.1136/adc.2002.024034 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
