

Regulation of Skeletal Muscle Function by Amino Acids
- PMID: 31963899
- PMCID: PMC7019684
- DOI: 10.3390/nu12010261
Regulation of Skeletal Muscle Function by Amino Acids
Abstract
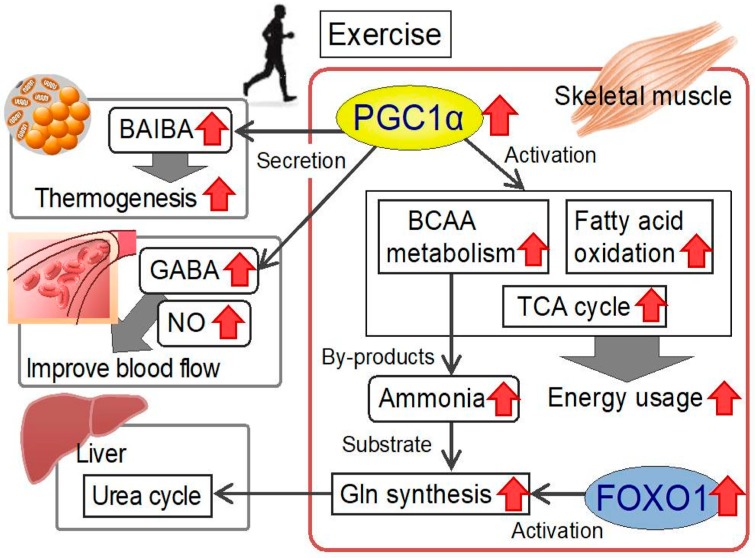
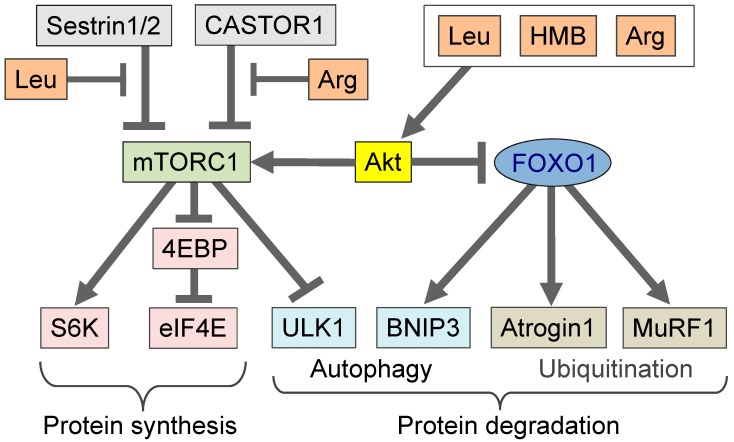
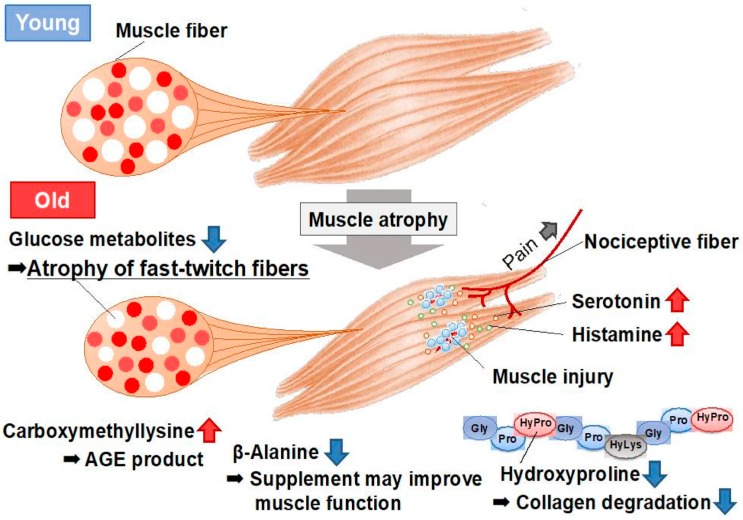
Amino acids are components of proteins that also exist free-form in the body; their functions can be divided into (1) nutritional, (2) sensory, and (3) biological regulatory roles. The skeletal muscle, which is the largest organ in the human body, representing ~40% of the total body weight, plays important roles in exercise, energy expenditure, and glucose/amino acid usage-processes that are modulated by various amino acids and their metabolites. In this review, we address the metabolism and function of amino acids in the skeletal muscle. The expression of PGC1α, a transcriptional coactivator, is increased in the skeletal muscle during exercise. PGC1α activates branched-chain amino acid (BCAA) metabolism and is used for energy in the tricarboxylic acid (TCA) cycle. Leucine, a BCAA, and its metabolite, β-hydroxy-β-methylbutyrate (HMB), both activate mammalian target of rapamycin complex 1 (mTORC1) and increase protein synthesis, but the mechanisms of activation appear to be different. The metabolite of valine (another BCAA), β-aminoisobutyric acid (BAIBA), is increased by exercise, is secreted by the skeletal muscle, and acts on other tissues, such as white adipose tissue, to increase energy expenditure. In addition, several amino acid-related molecules reportedly activate skeletal muscle function. Oral 5-aminolevulinic acid (ALA) supplementation can protect against mild hyperglycemia and help prevent type 2 diabetes. β-alanine levels are decreased in the skeletal muscles of aged mice. β-alanine supplementation increased the physical performance and improved the executive function induced by endurance exercise in middle-aged individuals. Further studies focusing on the effects of amino acids and their metabolites on skeletal muscle function will provide data essential for the production of food supplements for older adults, athletes, and individuals with metabolic diseases.
Keywords: PGC1α; amino acid; branched-chain amino acid (BCAA); energy expenditure; exercise; leucine; metabolic diseases; skeletal muscle; β-aminoisobutyric acid (BAIBA); β-hydroxy-β-methylbutyrate (HMB).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
