

Resource fluctuations inhibit the reproduction and virulence of the human parasite Schistosoma mansoni in its snail intermediate host
- PMID: 31964301
- PMCID: PMC7015343
- DOI: 10.1098/rspb.2019.2446
Resource fluctuations inhibit the reproduction and virulence of the human parasite Schistosoma mansoni in its snail intermediate host
Abstract
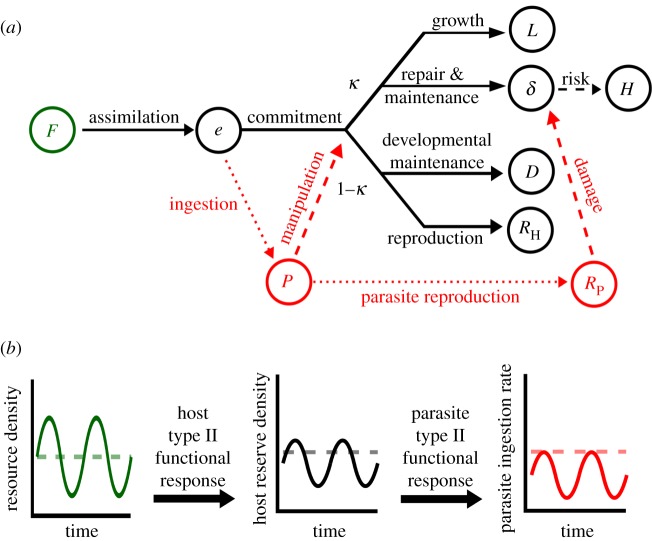
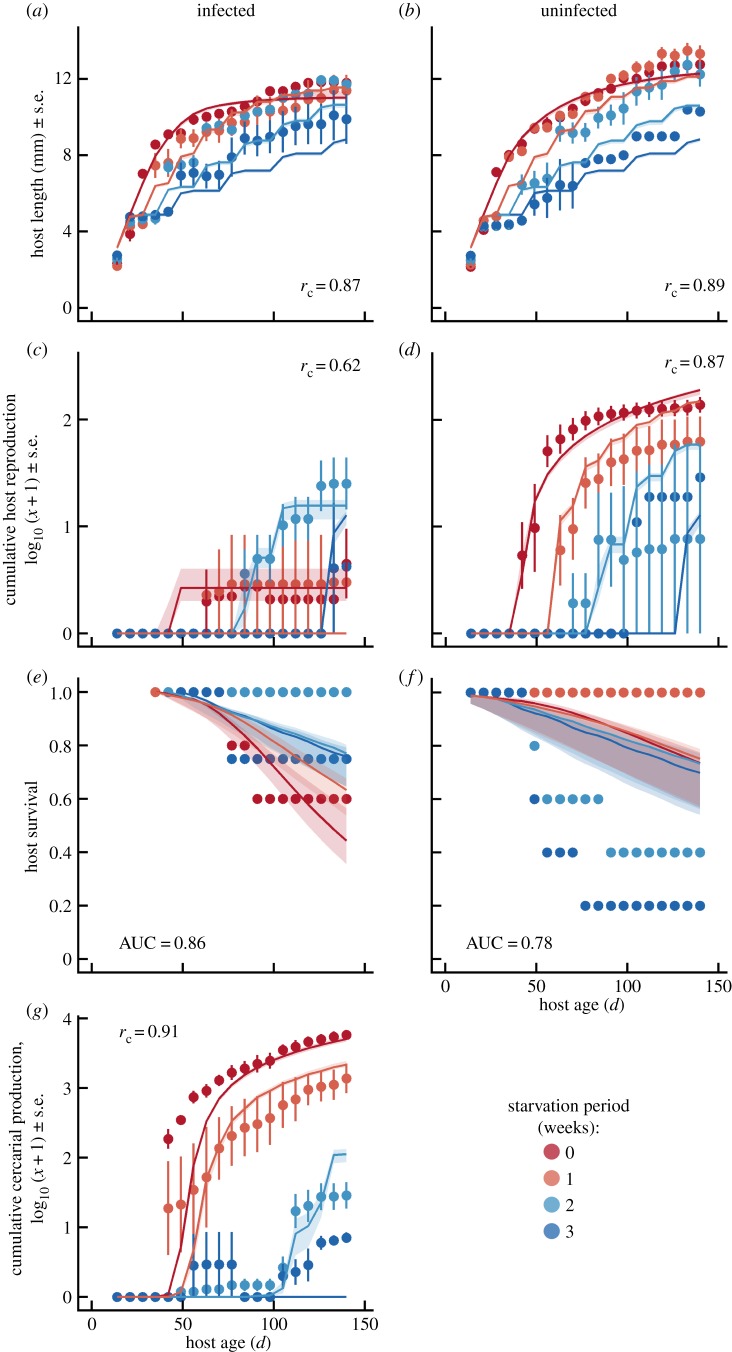
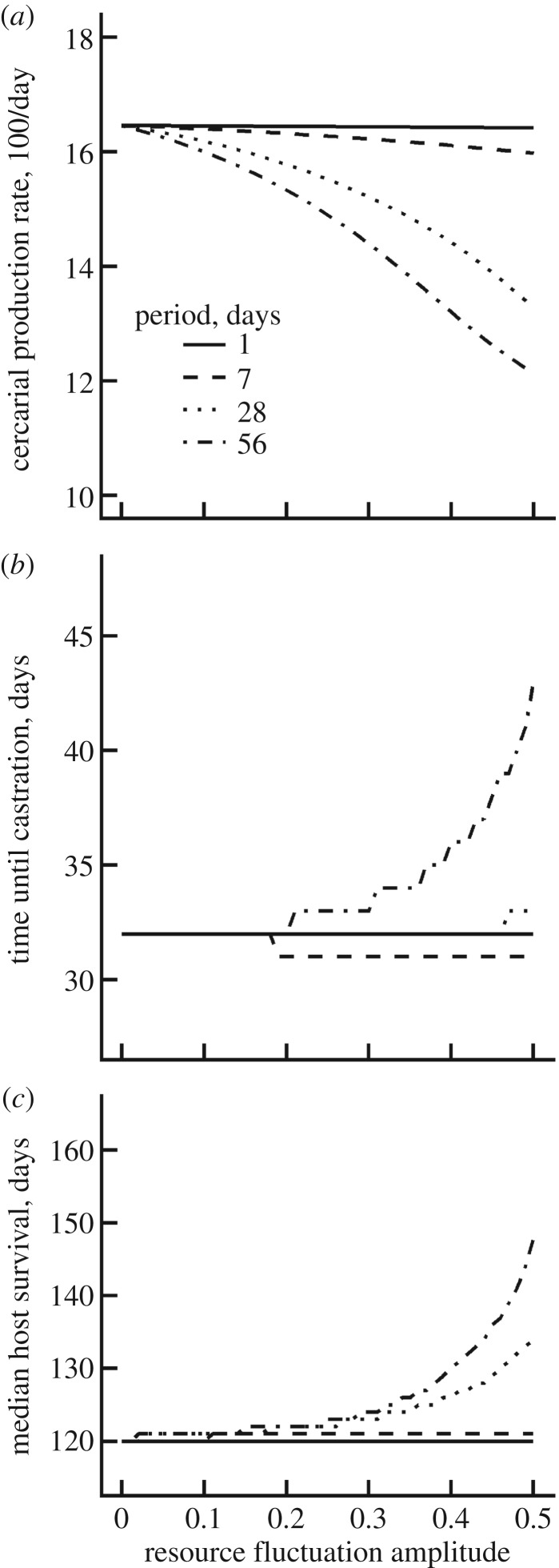
Resource availability can powerfully influence host-parasite interactions. However, we currently lack a mechanistic framework to predict how resource fluctuations alter individual infection dynamics. We address this gap with experiments manipulating resource supply and starvation for a human parasite, Schistosoma mansoni, and its snail intermediate host to test a hypothesis derived from mechanistic energy budget theory: resource fluctuations should reduce schistosome reproduction and virulence by inhibiting parasite ingestion of host biomass. Low resource supply caused hosts to remain small, reproduce less and produce fewer human-infectious cercariae. Periodic starvation also inhibited cercarial production and prevented infection-induced castration. The periodic starvation experiment also revealed substantial differences in fit between two bioenergetic model variants, which differ in their representation of host starvation. Simulations using the best-fit parameters of the winning model suggest that schistosome performance substantially declines with resource fluctuations with periods greater than 7 days. These experiments strengthen mechanistic theory, which can be readily scaled up to the population level to understand key feedbacks between resources, host population dynamics, parasitism and control interventions. Integrating resources with other environmental drivers of disease in an explicit bioenergetic framework could ultimately yield mechanistic predictions for many disease systems.
Keywords: energy budget; fluctuation; parasite production; parasitism; reproduction; resources.
Conflict of interest statement
We declare we have no competing interests.
Figures

References
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous