

A tripartite cooperative mechanism confers resistance of the protein kinase A catalytic subunit to dephosphorylation
- PMID: 31964716
- PMCID: PMC7062156
- DOI: 10.1074/jbc.RA119.010004
A tripartite cooperative mechanism confers resistance of the protein kinase A catalytic subunit to dephosphorylation
Abstract
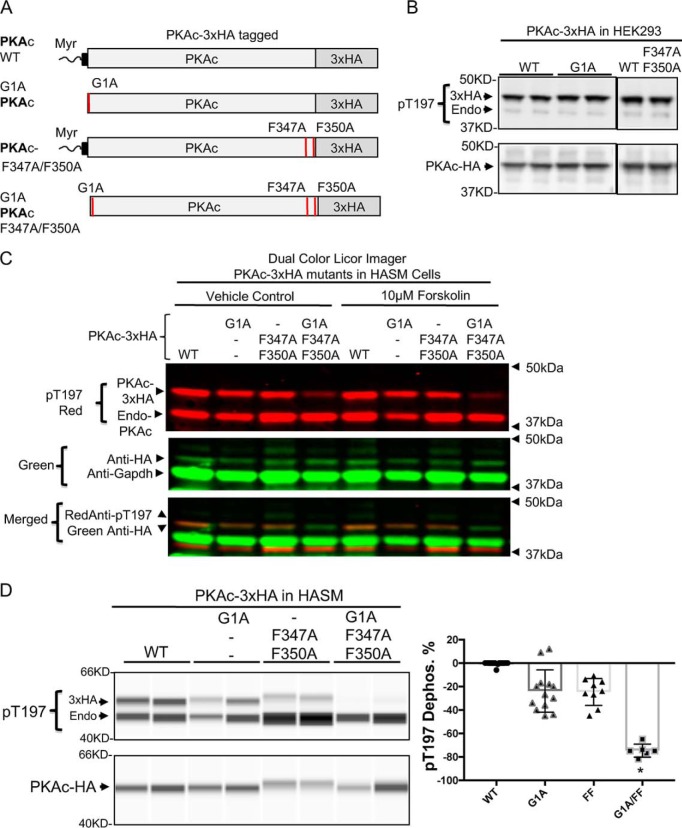
Phosphorylation of specific residues in the activation loops of AGC kinase group (protein kinase A, G, and C families) is required for activity of most of these kinases, including the catalytic subunit of PKA (PKAc). Although many phosphorylated AGC kinases are sensitive to phosphatase-mediated dephosphorylation, the PKAc activation loop uniquely resists dephosphorylation, rendering it "constitutively" phosphorylated in cells. Previous biophysical experiments and structural modeling have suggested that the N-terminal myristoylation signal and the C-terminal FXXF motif in PKAc regulate its thermal stability and catalysis. Here, using site-directed mutagenesis, molecular modeling, and in cell-free and cell-based systems, we demonstrate that substitutions of either the PKAc myristoylation signal or the FXXF motif only modestly reduce phosphorylation and fail to affect PKAc function in cells. However, we observed that these two sites cooperate with an N-terminal FXXW motif to cooperatively establish phosphatase resistance of PKAc while not affecting kinase-dependent phosphorylation of the activation loop. We noted that this tripartite cooperative mechanism of phosphatase resistance is functionally relevant, as demonstrated by changes in morphology, adhesion, and migration of human airway smooth muscle cells transfected with PKAc variants containing amino acid substitutions in these three sites. These findings establish that three allosteric sites located at the PKAc N and C termini coordinately regulate the phosphatase sensitivity of this enzyme. This cooperative mechanism of phosphatase resistance of AGC kinase opens new perspectives toward therapeutic manipulation of kinase signaling in disease.
Keywords: Akt PKB; PKA; cAMP-dependent protein kinase; dephosphorylation; phosphatase; protein kinase B; serine/threonine protein kinase; signal transduction; structure–function.
© 2020 Chan et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health
Figures





References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
