

IFITM3 Reduces Retroviral Envelope Abundance and Function and Is Counteracted by glycoGag
- PMID: 31964738
- PMCID: PMC6974572
- DOI: 10.1128/mBio.03088-19
IFITM3 Reduces Retroviral Envelope Abundance and Function and Is Counteracted by glycoGag
Abstract
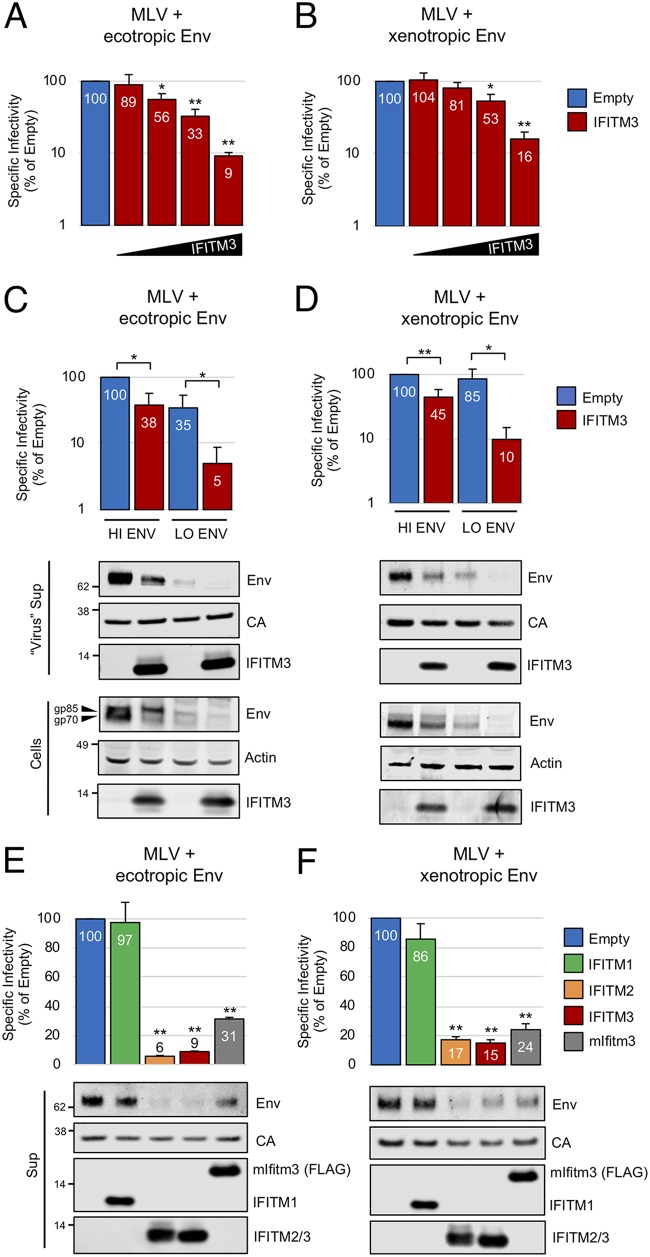
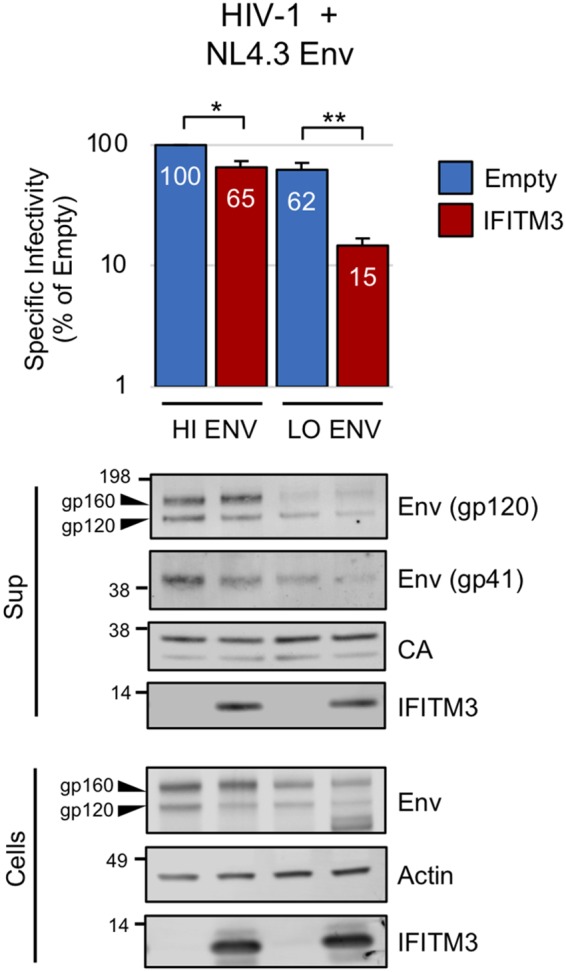
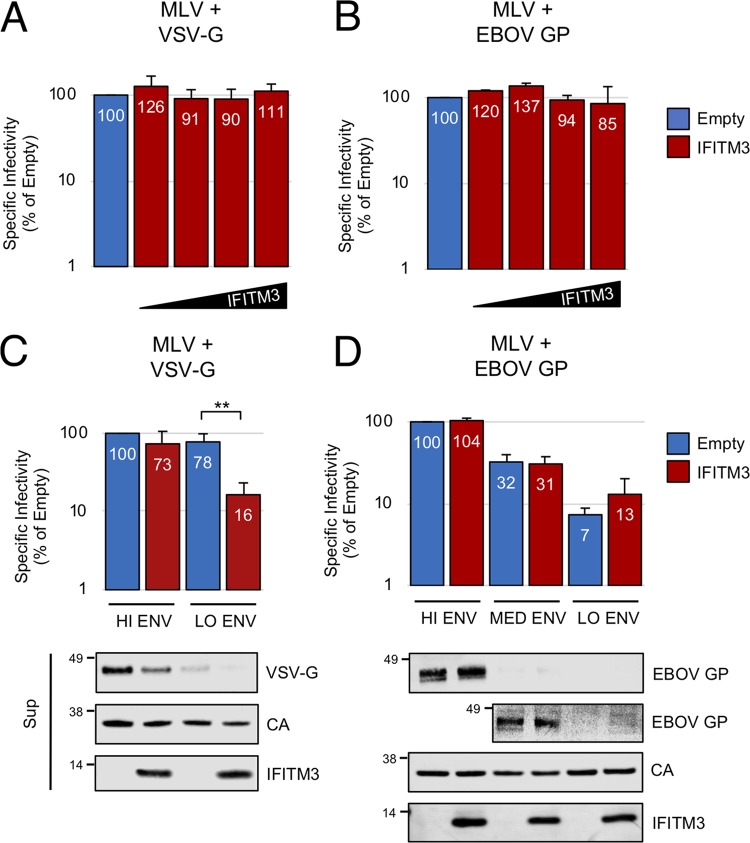
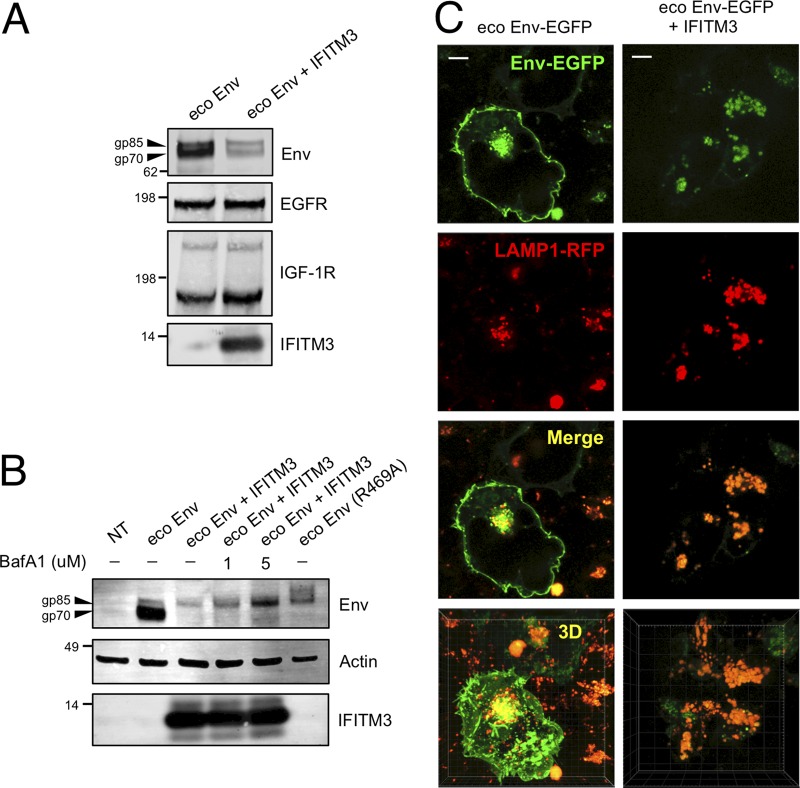
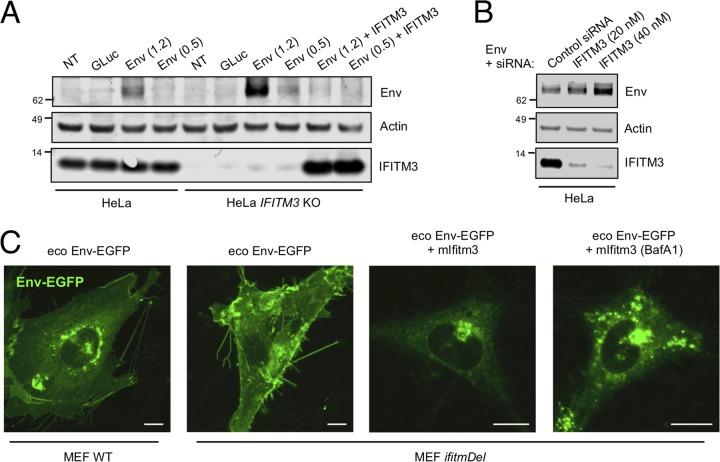
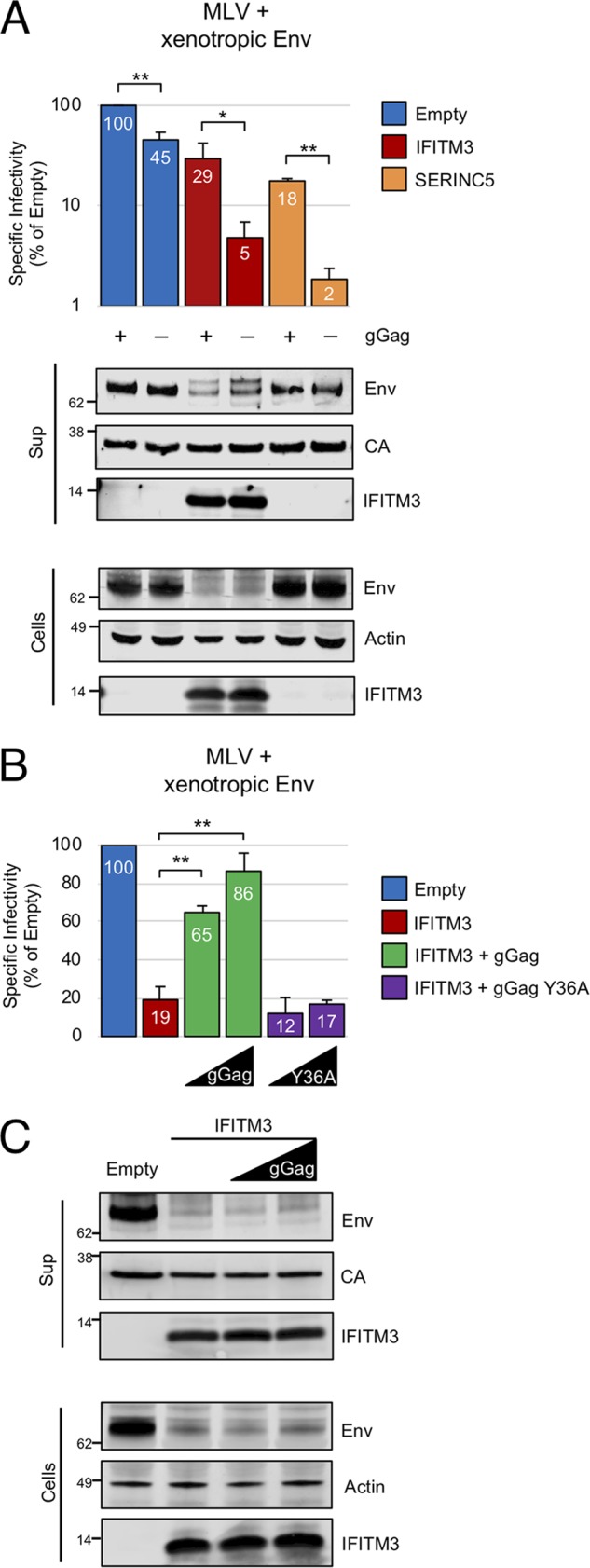
Interferon-induced transmembrane (IFITM) proteins are encoded by many vertebrate species and exhibit antiviral activities against a wide range of viruses. IFITM3, when present in virus-producing cells, reduces the fusion potential of HIV-1 virions, but the mechanism is poorly understood. To define the breadth and mechanistic basis for the antiviral activity of IFITM3, we took advantage of a murine leukemia virus (MLV)-based pseudotyping system. By carefully controlling amounts of IFITM3 and envelope protein (Env) in virus-producing cells, we found that IFITM3 potently inhibits MLV infectivity when Env levels are limiting. Loss of infectivity was associated with defective proteolytic processing of Env and lysosomal degradation of the Env precursor. Ecotropic and xenotropic variants of MLV Env, as well as HIV-1 Env and vesicular stomatitis virus glycoprotein (VSV-G), are sensitive to IFITM3, whereas Ebola glycoprotein is resistant, suggesting that IFITM3 selectively inactivates certain viral glycoproteins. Furthermore, endogenous IFITM3 in human and murine cells negatively regulates MLV Env abundance. However, we found that the negative impact of IFITM3 on virion infectivity is greater than its impact on decreasing Env incorporation, suggesting that IFITM3 may impair Env function, as well as reduce the amount of Env in virions. Finally, we demonstrate that loss of virion infectivity mediated by IFITM3 is reversed by the expression of glycoGag, a murine retrovirus accessory protein previously shown to antagonize the antiviral activity of SERINC proteins. Overall, we show that IFITM3 impairs virion infectivity by regulating Env quantity and function but that enhanced Env expression and glycoGag confer viral resistance to IFITM3.IMPORTANCE The viral envelope glycoprotein, known as "Env" in Retroviridae, is found on the virion surface and facilitates virus entry into cells by mediating cell attachment and fusion. Env is a major structural component of retroviruses and is targeted by all arms of the immune response, including adaptive and innate immunity. Less is known about how cell-intrinsic immunity prevents retrovirus replication at the level of individual cells. Here, we show that cellular IFITM3 and IFITM2 inhibit the fusion potential of retroviral virions by inhibiting Env protein via a two-pronged mechanism. IFITM proteins inhibit Env abundance in cells and also impair its function when levels are low. The posttranslational block of retroviral Env function by IFITM proteins is likely to impede both exogenous and endogenous retrovirus replication. In support of a relevant role for IFITM3 in retrovirus control, the retroviral accessory protein glycoGag counteracts IFITM3 function to promote virus infectivity.
Keywords: Env; IFITM; fusion; innate immunity; retroviruses; viral glycoprotein.
Figures
References
-
- Braun E, Hotter D, Koepke L, Zech F, Gross R, Sparrer KMJ, Müller JA, Pfaller CK, Heusinger E, Wombacher R, Sutter K, Dittmer U, Winkler M, Simmons G, Jakobsen MR, Conzelmann K-K, Pöhlmann S, Münch J, Fackler OT, Kirchhoff F, Sauter D. 2019. Guanylate-binding proteins 2 and 5 exert broad antiviral activity by inhibiting furin-mediated processing of viral Envelope proteins. Cell Rep 27:2092–2104.e2010. doi:10.1016/j.celrep.2019.04.063. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
