

Nucleosomal proofreading of activator-promoter interactions
- PMID: 31964832
- PMCID: PMC7007518
- DOI: 10.1073/pnas.1911188117
Nucleosomal proofreading of activator-promoter interactions
Abstract
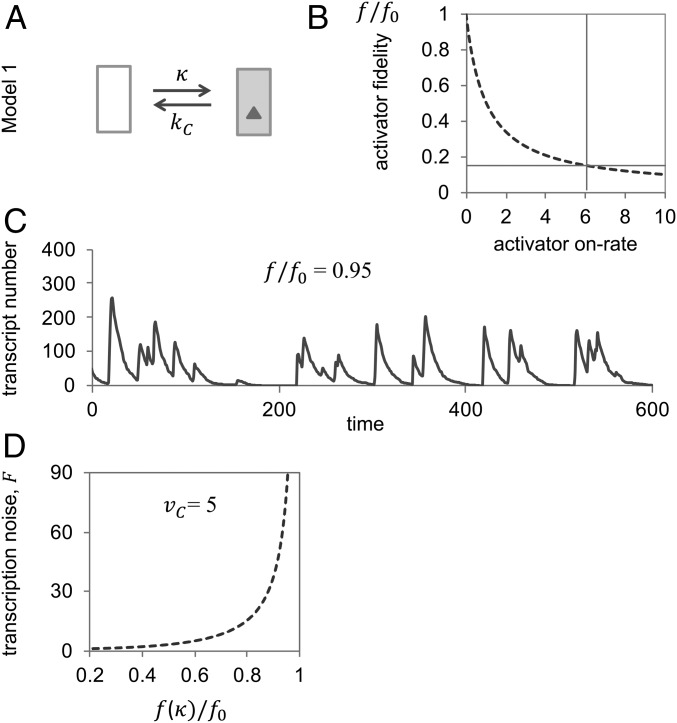
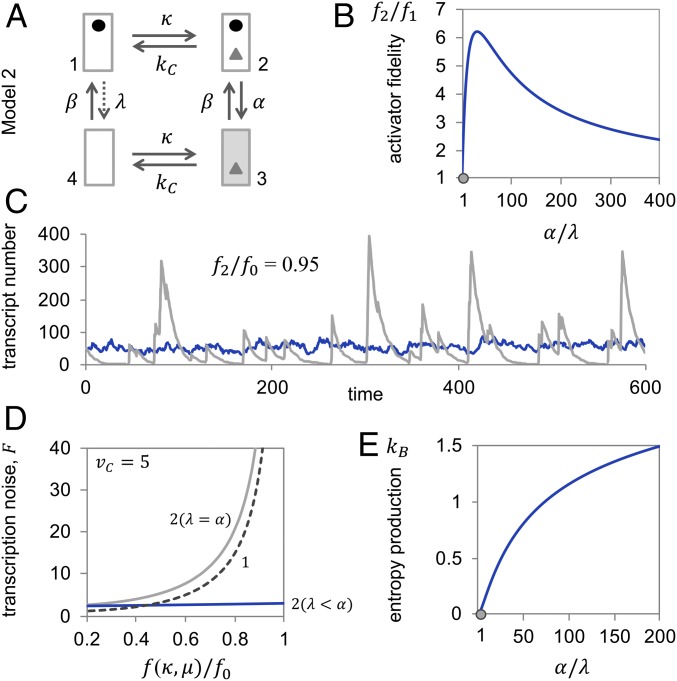
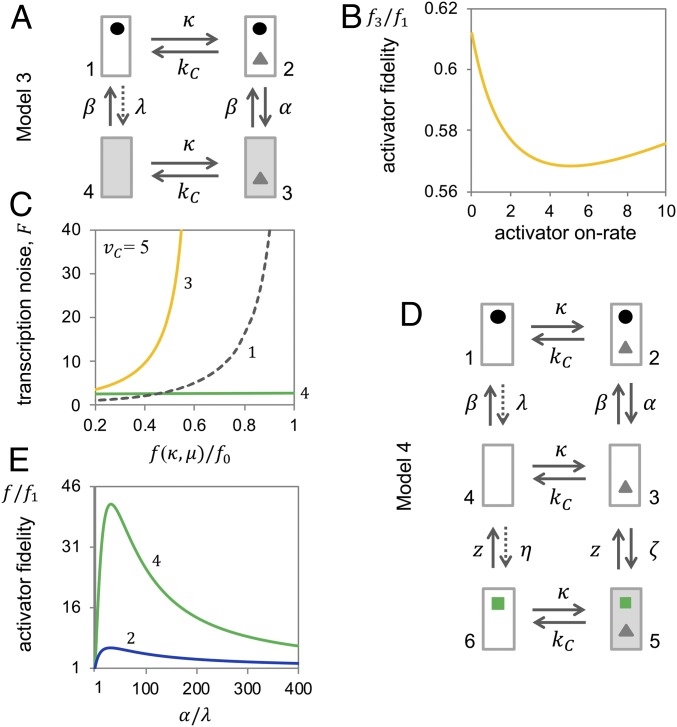
Specificity in transcriptional regulation is imparted by transcriptional activators that bind to specific DNA sequences from which they stimulate transcription. Specificity may be increased by slowing down the kinetics of regulation: by increasing the energy for dissociation of the activator-DNA complex or decreasing activator concentration. In general, higher dissociation energies imply longer DNA dwell times of the activator; the activator-bound gene may not readily turn off again. Lower activator concentrations entail longer pauses between binding events; the activator-unbound gene is not easily turned on again and activated transcription occurs in stochastic bursts. We show that kinetic proofreading of activator-DNA recognition-insertion of an energy-dissipating delay step into the activation pathway for transcription-reconciles high specificity of transcriptional regulation with fast regulatory kinetics. We show that kinetic proofreading results from the stochastic removal and reformation of promoter nucleosomes, at a distance from equilibrium.
Keywords: entropy production; irreversibility; kinetic proofreading; nucleosome; transcriptional regulation.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Ptashne M., Gann A., Transcriptional activation by recruitment. Nature 386, 569–577 (1997). - PubMed
-
- Bryant G. O., Ptashne M., Independent recruitment in vivo by Gal4 of two complexes required for transcription. Mol. Cell 11, 1301–1309 (2003). - PubMed
-
- Cosma M. P., Tanaka T., Nasmyth K., Ordered recruitment of transcription and chromatin remodeling factors to a cell cycle- and developmentally regulated promoter. Cell 97, 299–311 (1999). - PubMed
