

TET methylcytosine oxidases: new insights from a decade of research
- PMID: 31965999
- PMCID: PMC7216820
TET methylcytosine oxidases: new insights from a decade of research
Abstract
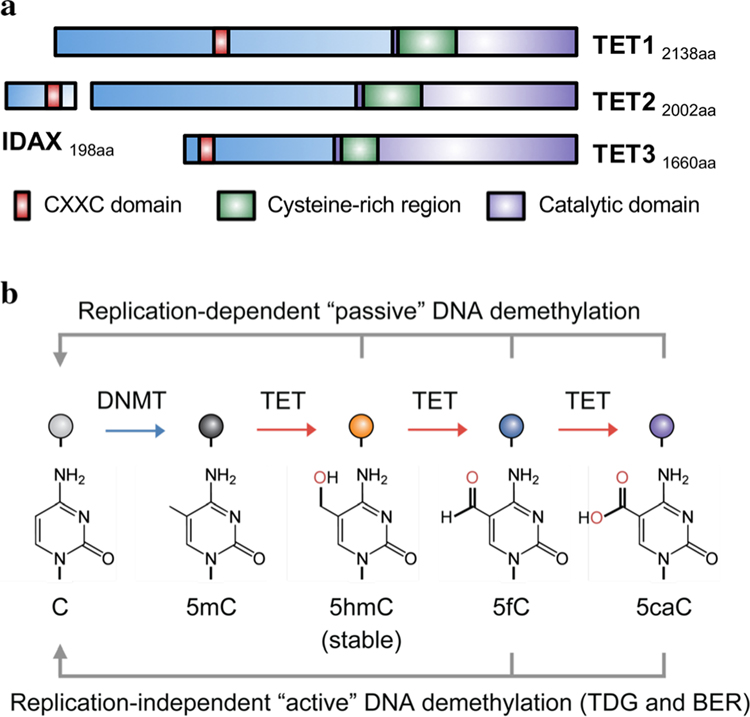
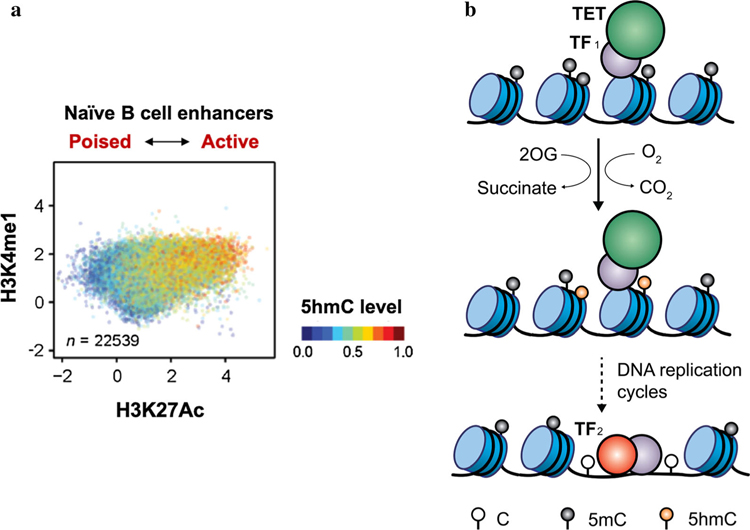
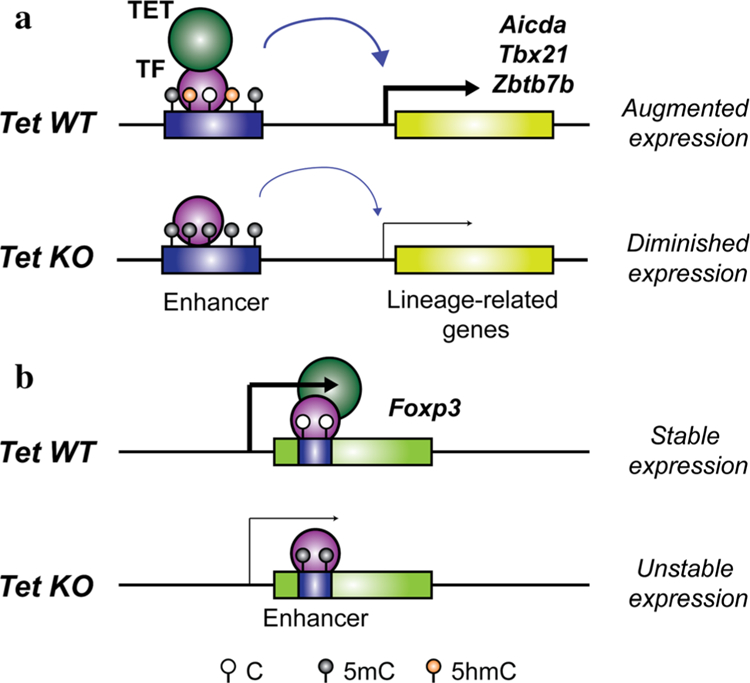
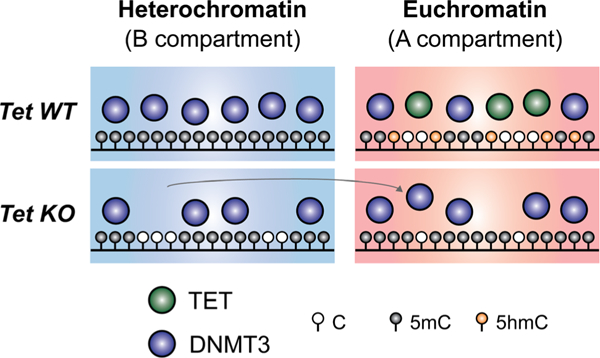
In mammals, DNA methyltransferases transfer a methyl group from S-adenosylmethionine to the 5 position of cytosine in DNA. The product of this reaction, 5-methylcytosine (5mC), has many roles, particularly in suppressing transposable and repeat elements in DNA. Moreover, in many cellular systems, cell lineage specification is accompanied by DNA demethylation at the promoters of genes expressed at high levels in the differentiated cells. However, since direct cleavage of the C-C bond connecting the methyl group to the 5 position of cytosine is thermodynamically disfavoured, the question of whether DNA methylation was reversible remained unclear for many decades. This puzzle was solved by our discovery of the TET (Ten- Eleven Translocation) family of 5-methylcytosine oxidases, which use reduced iron, molecular oxygen and the tricarboxylic acid cycle metabolite 2-oxoglutarate (also known as a-ketoglutarate) to oxidise the methyl group of 5mC to 5-hydroxymethylcytosine (5hmC) and beyond. TET-generated oxidised methylcytosines are intermediates in at least two pathways of DNA demethylation, which differ in their dependence on DNA replication. In the decade since their discovery, TET enzymes have been shown to have important roles in embryonic development, cell lineage specification, neuronal function and cancer. We review these findings and discuss their implications here.
Conflict of interest statement
Figures
References
-
- Aravind L, Abhiman S, and Iyer LM 2011. Natural history of the eukaryotic chromatin protein methylation system. Prog Mol Biol Transl Sci 101 105–176 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
