

New Roles for the Heterochronic Transcription Factor LIN-29 in Cuticle Maintenance and Lipid Metabolism at the Larval-to-Adult Transition in Caenorhabditis elegans
- PMID: 31974205
- PMCID: PMC7054012
- DOI: 10.1534/genetics.119.302860
New Roles for the Heterochronic Transcription Factor LIN-29 in Cuticle Maintenance and Lipid Metabolism at the Larval-to-Adult Transition in Caenorhabditis elegans
Abstract
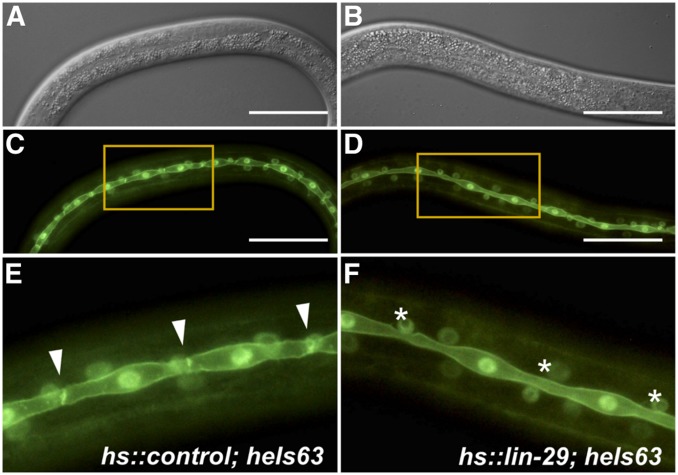
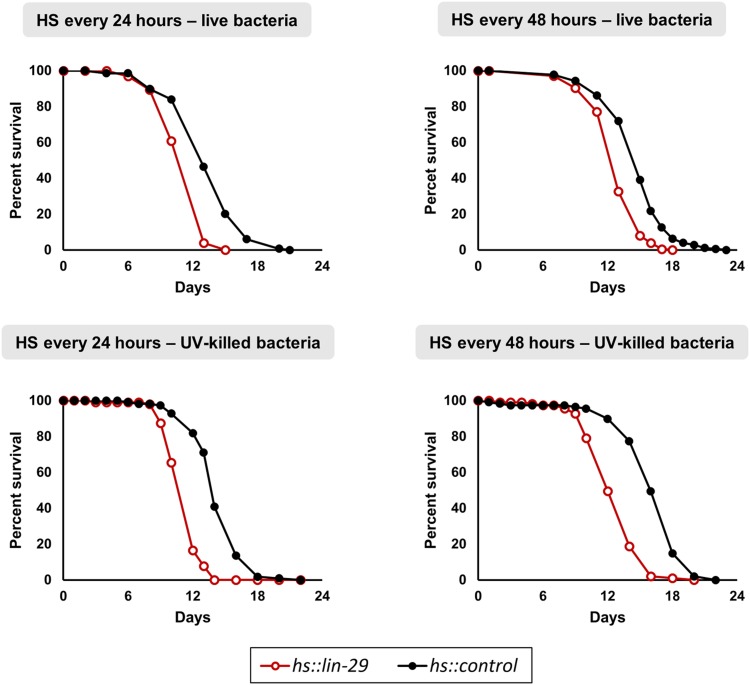
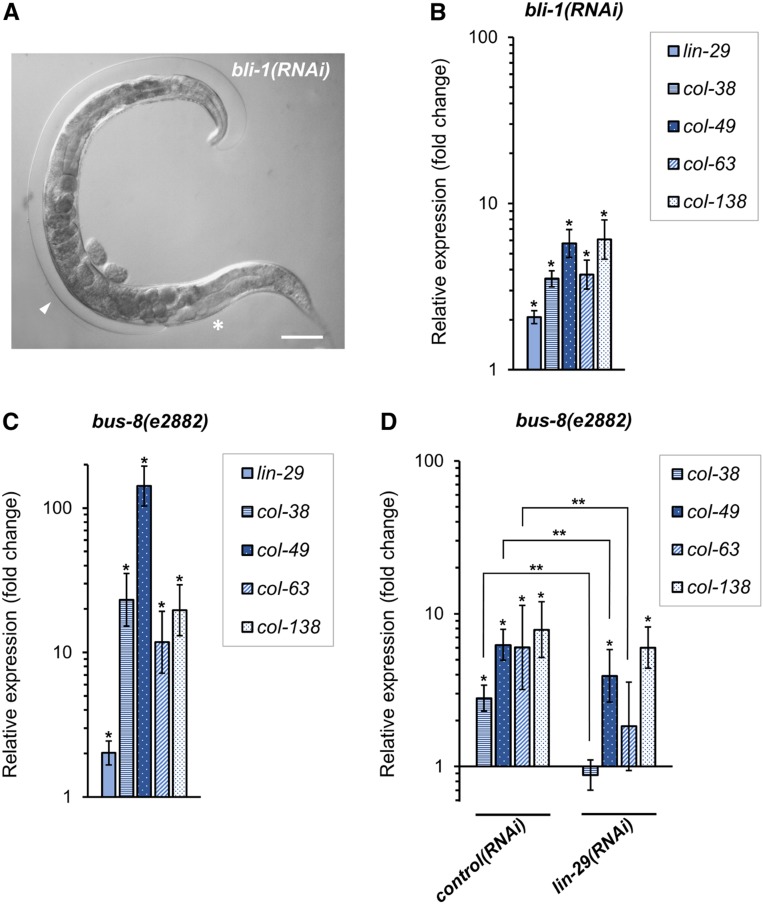
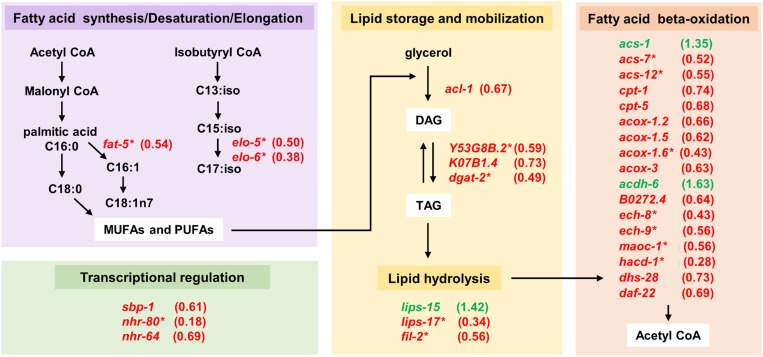
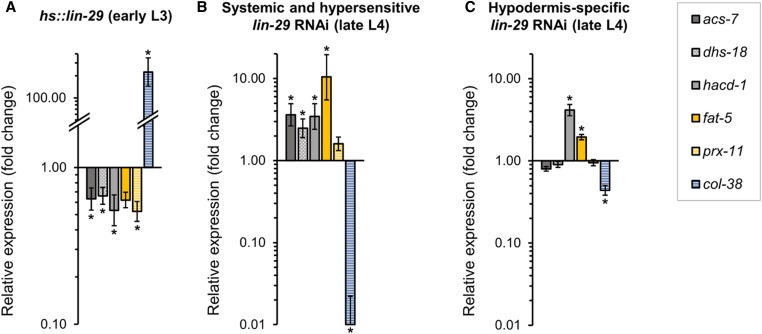
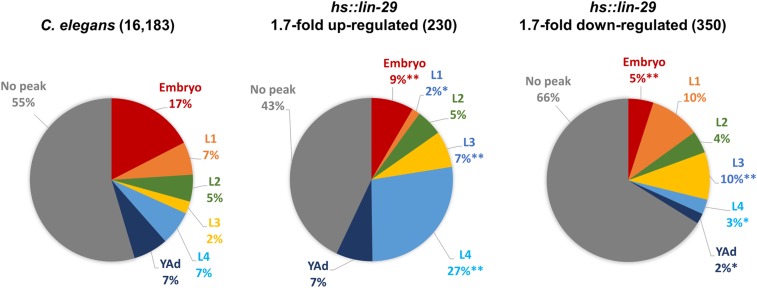
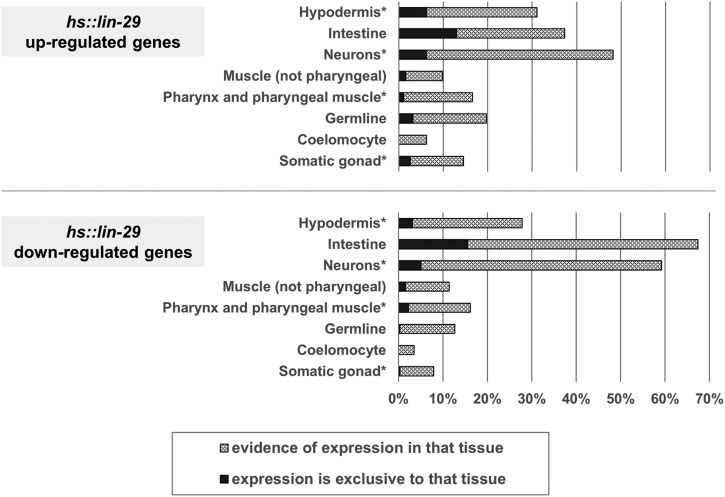
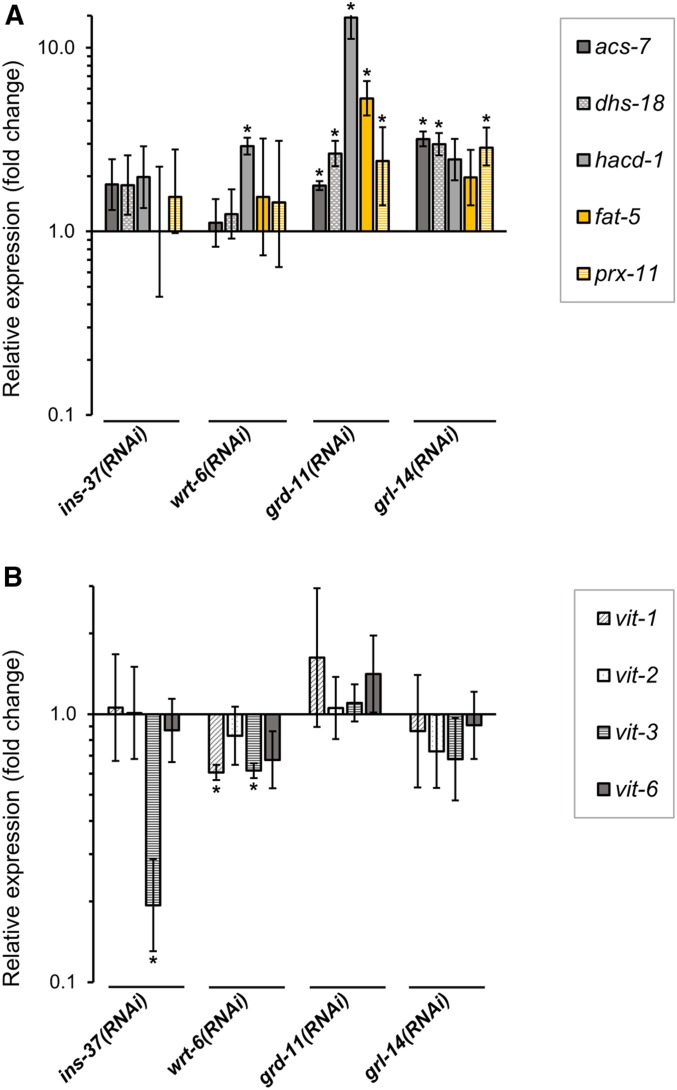
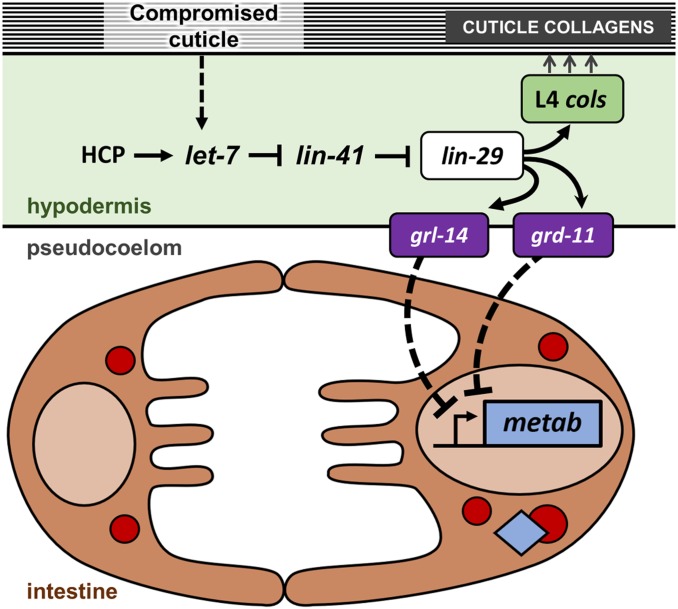
Temporal regulation of gene expression is a crucial aspect of metazoan development. In the roundworm Caenorhabditis elegans, the heterochronic pathway controls multiple developmental events in a time-specific manner. The most downstream effector of this pathway, the zinc-finger transcription factor LIN-29, acts in the last larval stage (L4) to regulate elements of the larval-to-adult switch. Here, we explore new LIN-29 targets and their implications for this developmental transition. We used RNA-sequencing to identify genes differentially expressed between animals misexpressing LIN-29 at an early time point and control animals. Among 230 LIN-29-activated genes, we found that genes encoding cuticle collagens were overrepresented. Interestingly, expression of lin-29 and some of these collagens was increased in adults with cuticle damage, suggesting a previously unknown function for LIN-29 in adult cuticle maintenance. On the other hand, genes involved in fat metabolism were enriched among 350 LIN-29-downregulated targets. We used mass spectrometry to assay lipid content in animals overexpressing LIN-29 and observed reduced fatty acid levels. Many LIN-29-repressed genes are normally expressed in the intestine, suggesting cell-nonautonomous regulation. We identified several LIN-29 upregulated genes encoding signaling molecules that may act as mediators in the regulation of intestinally expressed genes encoding fat metabolic enzymes and vitellogenins. Overall, our results support the model of LIN-29 as a major regulator of adult cuticle synthesis and integrity, and as the trigger for metabolic changes that take place at the important transition from rapid growth during larval life to slower growth and offspring production during adulthood.
Keywords: Caenorhabditis elegans; collagen; gene expression; heterochronic; metabolism.
Copyright © 2020 by the Genetics Society of America.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
