

Mycelial network-mediated rhizobial dispersal enhances legume nodulation
- PMID: 31974462
- PMCID: PMC7082348
- DOI: 10.1038/s41396-020-0587-5
Mycelial network-mediated rhizobial dispersal enhances legume nodulation
Abstract
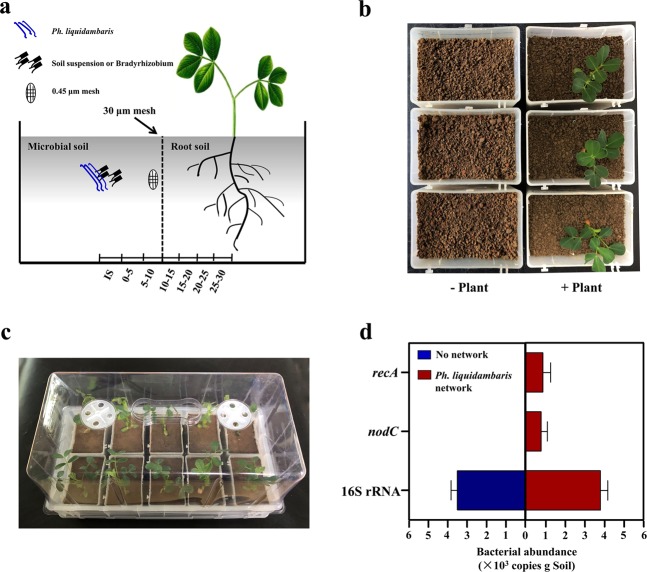
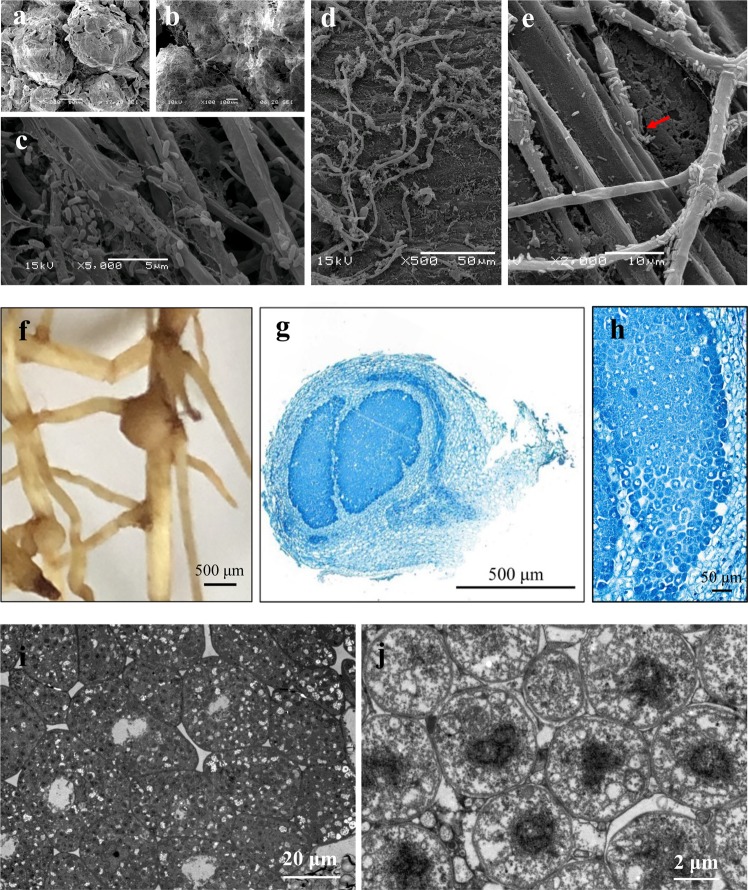
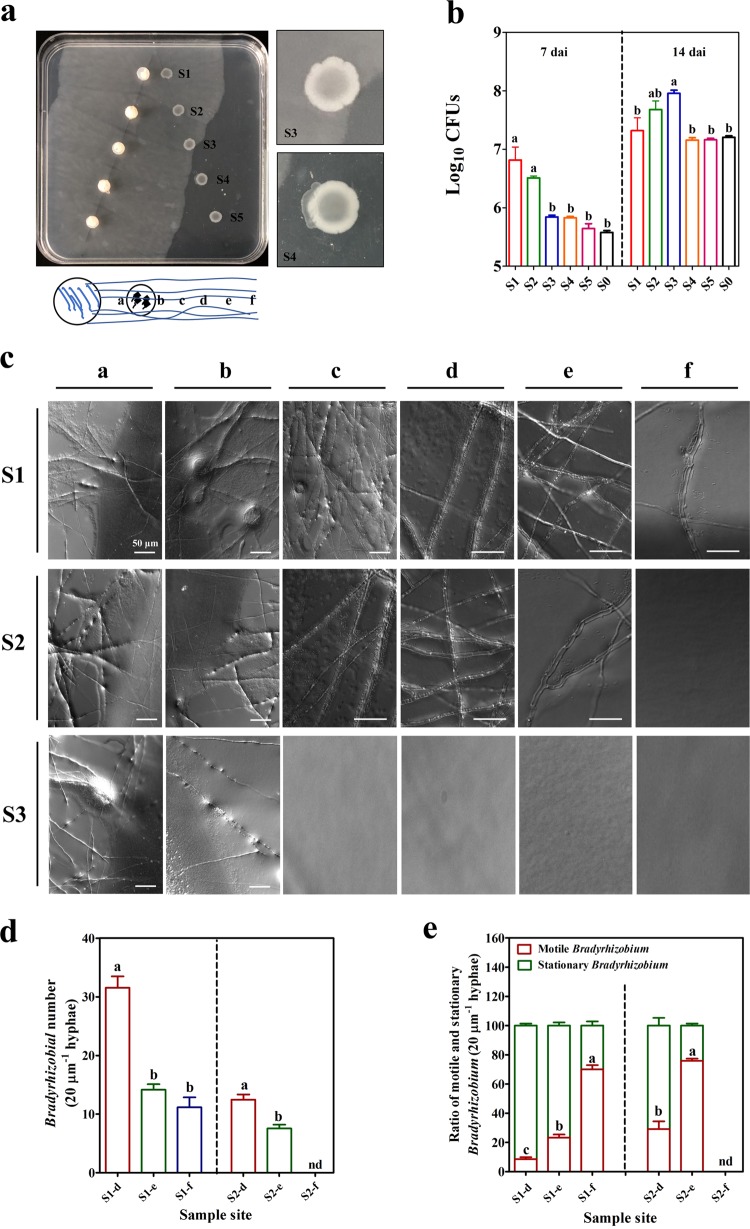
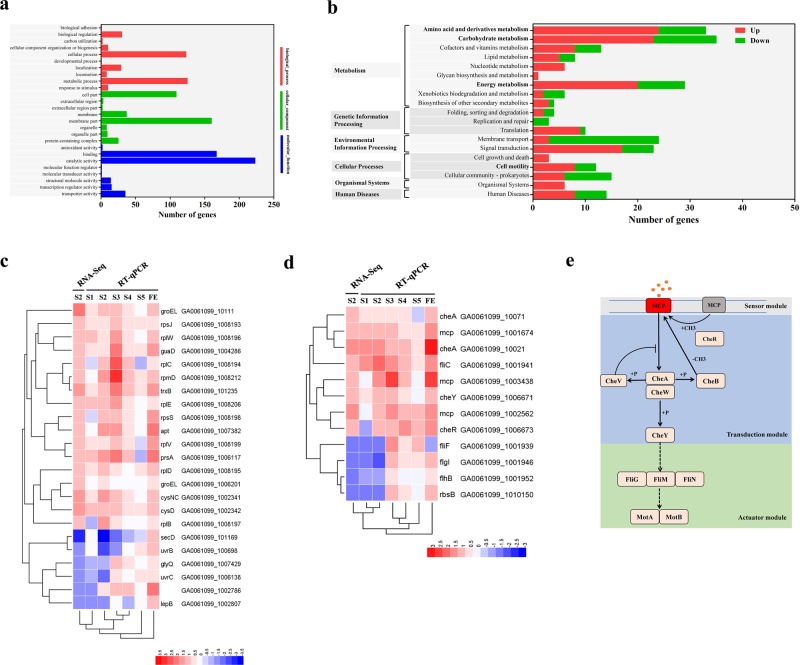
The access of rhizobia to legume host is a prerequisite for nodulation. Rhizobia are poorly motile in soil, while filamentous fungi are known to grow extensively across soil pores. Since root exudates-driven bacterial chemotaxis cannot explain rhizobial long-distance dispersal, mycelia could constitute ideal dispersal networks to help rhizobial enrichment in the legume rhizosphere from bulk soil. Thus, we hypothesized that mycelia networks act as vectors that enable contact between rhizobia and legume and influence subsequent nodulation. By developing a soil microcosm system, we found that a facultatively biotrophic fungus, Phomopsis liquidambaris, helps rhizobial migration from bulk soil to the peanut (Arachis hypogaea) rhizosphere and, hence, triggers peanut-rhizobium nodulation but not seen in the absence of mycelia. Assays of dispersal modes suggested that cell proliferation and motility mediated rhizobial dispersal along mycelia, and fungal exudates might contribute to this process. Furthermore, transcriptomic analysis indicated that genes associated with the cell division, chemosensory system, flagellum biosynthesis, and motility were regulated by Ph. liquidambaris, thus accounting for the detected rhizobial dispersal along hyphae. Our results indicate that rhizobia use mycelia as dispersal networks that migrate to legume rhizosphere and trigger nodulation. This work highlights the importance of mycelial network-based bacterial dispersal in legume-rhizobium symbiosis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



Similar articles
-
Arbuscular mycorrhizal hyphae facilitate rhizobia dispersal and nodulation in legumes.ISME J. 2024 Jan 8;18(1):wrae185. doi: 10.1093/ismejo/wrae185. ISME J. 2024. PMID: 39325968 Free PMC article.
-
Priming of rhizobial nodulation signaling in the mycosphere accelerates nodulation of legume hosts.New Phytol. 2022 Aug;235(3):1212-1230. doi: 10.1111/nph.18192. Epub 2022 May 23. New Phytol. 2022. PMID: 35488499
-
Endophytic Fungus Drives Nodulation and N2 Fixation Attributable to Specific Root Exudates.mBio. 2019 Jul 16;10(4):e00728-19. doi: 10.1128/mBio.00728-19. mBio. 2019. PMID: 31311876 Free PMC article.
-
Coevolution in Rhizobium-legume symbiosis?DNA Cell Biol. 2009 Aug;28(8):361-70. doi: 10.1089/dna.2009.0863. DNA Cell Biol. 2009. PMID: 19485766 Review.
-
Challenges to rhizobial adaptability in a changing climate: Genetic engineering solutions for stress tolerance.Microbiol Res. 2024 Nov;288:127886. doi: 10.1016/j.micres.2024.127886. Epub 2024 Aug 31. Microbiol Res. 2024. PMID: 39232483 Review.
Cited by
-
Population Response of Rhizosphere Microbiota of Garden Pea Genotypes to Inoculation with Arbuscular Mycorrhizal Fungi.Int J Mol Sci. 2023 Jan 6;24(2):1119. doi: 10.3390/ijms24021119. Int J Mol Sci. 2023. PMID: 36674632 Free PMC article.
-
Biological and experimental factors that define the effectiveness of microbial inoculation on plant traits: a meta-analysis.ISME Commun. 2024 Oct 14;4(1):ycae122. doi: 10.1093/ismeco/ycae122. eCollection 2024 Jan. ISME Commun. 2024. PMID: 39507396 Free PMC article.
-
The disruption of the MAPKK gene triggering the synthesis of flavonoids in endophytic fungus Phomopsis liquidambaris.Biotechnol Lett. 2021 Jan;43(1):119-132. doi: 10.1007/s10529-020-03042-5. Epub 2020 Oct 31. Biotechnol Lett. 2021. PMID: 33128663
-
Proximity-based defensive mutualism between Streptomyces and Mesorhizobium by sharing and sequestering iron.ISME J. 2024 Jan 8;18(1):wrad041. doi: 10.1093/ismejo/wrad041. ISME J. 2024. PMID: 38366066 Free PMC article.
-
Arbuscular mycorrhizal hyphae facilitate rhizobia dispersal and nodulation in legumes.ISME J. 2024 Jan 8;18(1):wrae185. doi: 10.1093/ismejo/wrae185. ISME J. 2024. PMID: 39325968 Free PMC article.
References
-
- Zipfel C, Oldroyd GED. Plant signalling in symbiosis and immunity. Nature. 2017;543:328–36. - PubMed
-
- Burchill W, James EK, Li D, Lanigan GJ, Williams M, Iannetta PPM, et al. Comparisons of biological nitrogen fixation in association with white clover (Trifolium repens L.) under four fertilizer nitrogen inputs as measured using two 15N techniques. Plant Soil. 2014;385:287–302.
-
- López-García SL, Perticari A, Piccinetti C, Ventimiglia L, Arias N, De Battista JJ, et al. In-furrow inoculation and selection for higher motility enhances the efficacy of Bradyrhizobium japonicum nodulation. Agron J. 2009;101:357–63.
-
- Vicario JC, Dardanelli MS, Giordano W. Swimming and swarming motility properties of peanut-nodulating rhizobia. FEMS Microbiol Lett. 2015;362:1–6. - PubMed
