

Optogenetics reveals Cdc42 local activation by scaffold-mediated positive feedback and Ras GTPase
- PMID: 31978045
- PMCID: PMC7002011
- DOI: 10.1371/journal.pbio.3000600
Optogenetics reveals Cdc42 local activation by scaffold-mediated positive feedback and Ras GTPase
Abstract
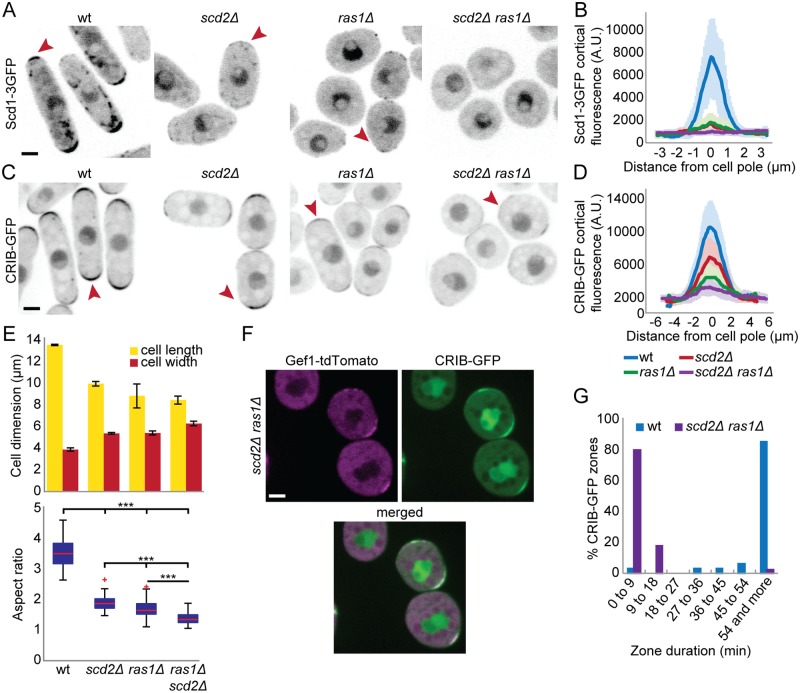
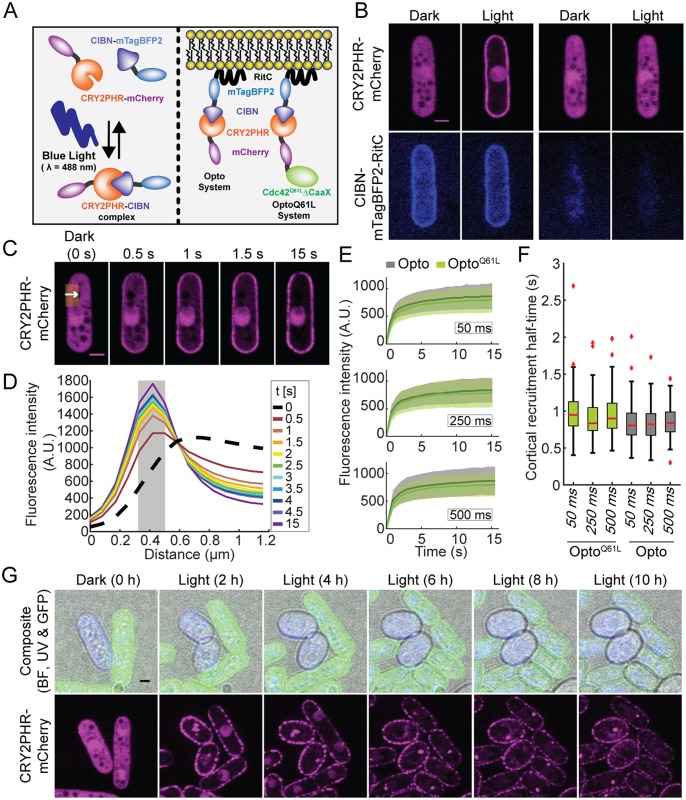
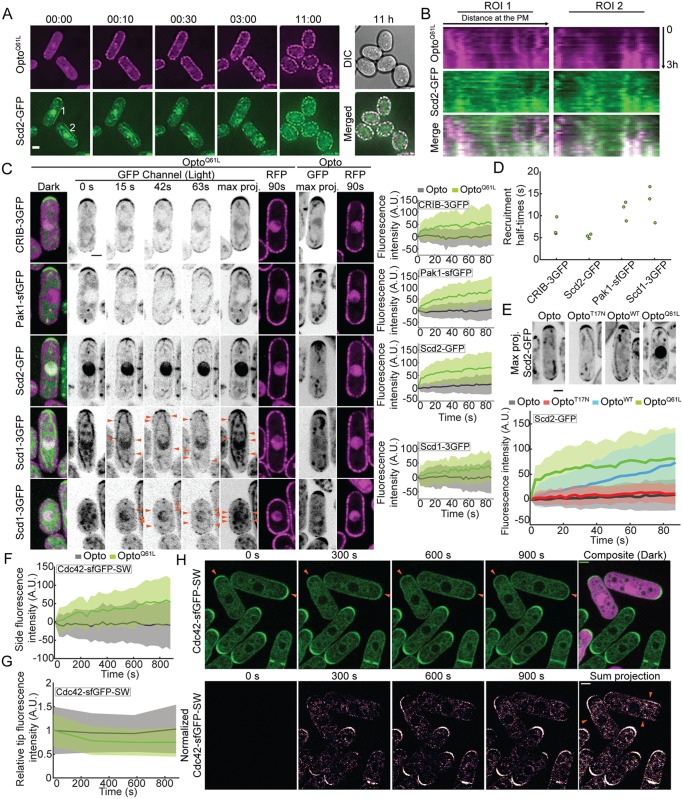
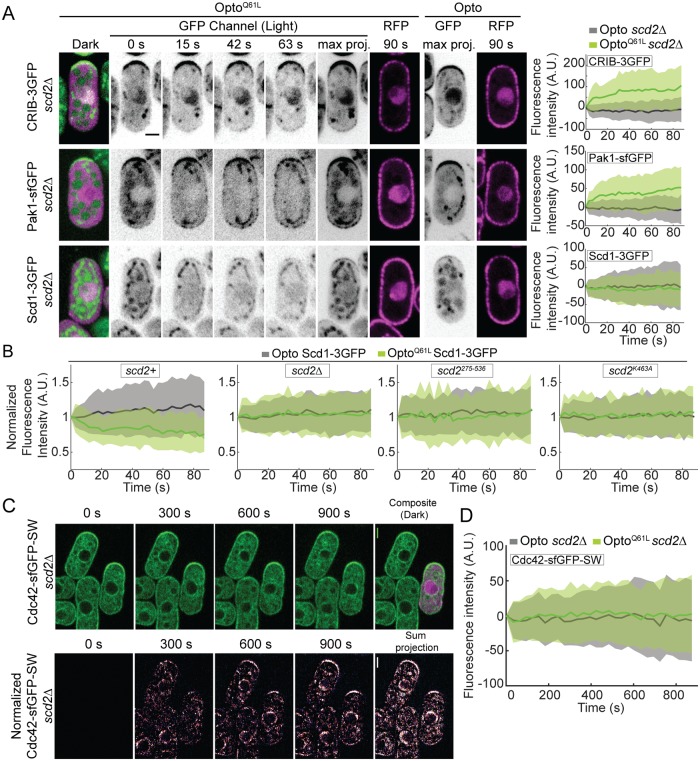
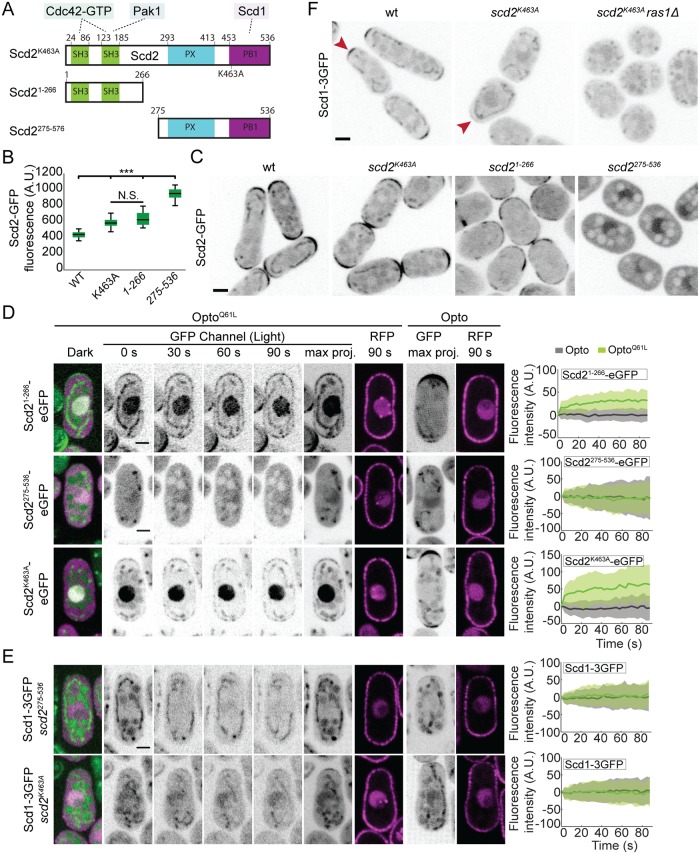
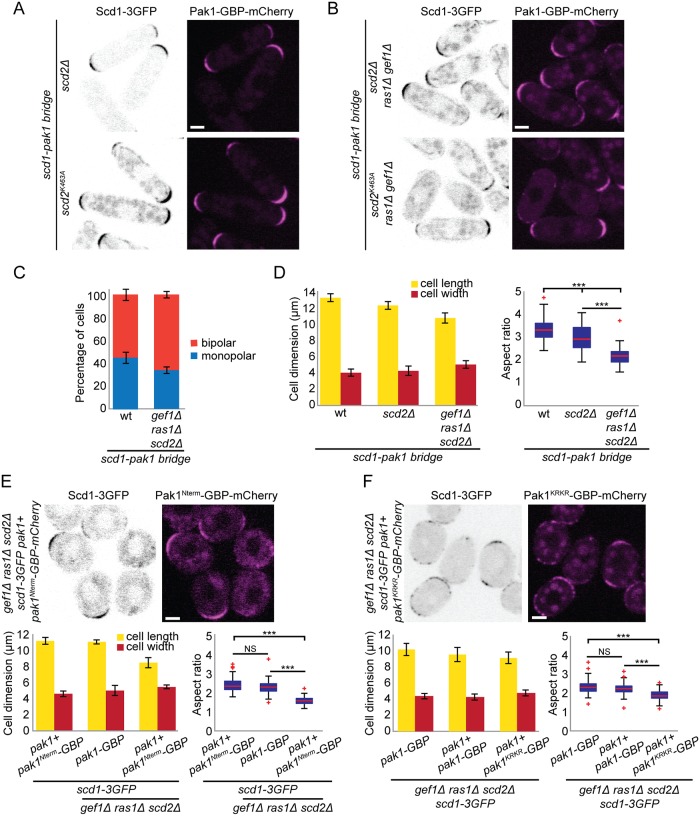
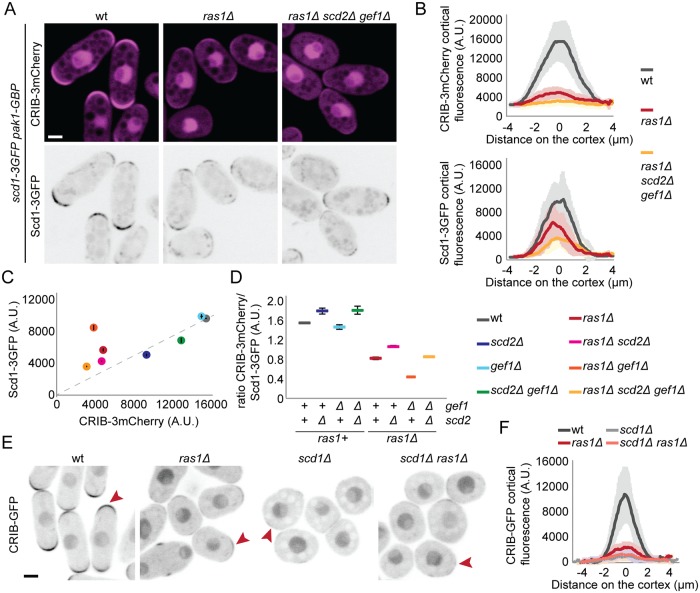
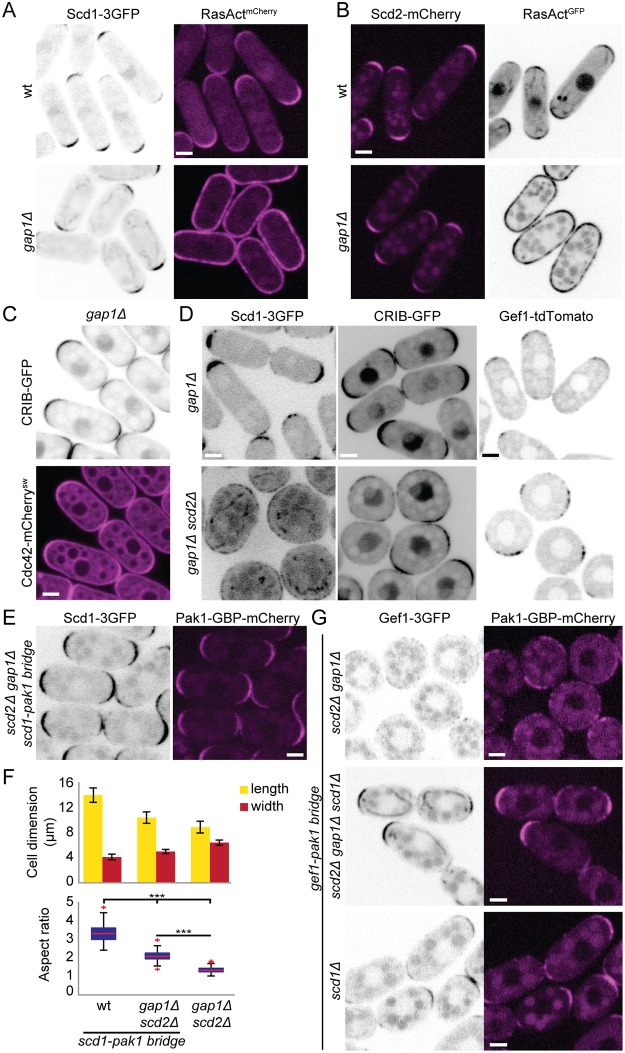
Local activity of the small GTPase Cdc42 is critical for cell polarization. Whereas scaffold-mediated positive feedback was proposed to break symmetry of budding yeast cells and produce a single zone of Cdc42 activity, the existence of similar regulation has not been probed in other organisms. Here, we address this problem using rod-shaped cells of fission yeast Schizosaccharomyces pombe, which exhibit zones of active Cdc42-GTP at both cell poles. We implemented the CRY2-CIB1 optogenetic system for acute light-dependent protein recruitment to the plasma membrane, which allowed to directly demonstrate positive feedback. Indeed, optogenetic recruitment of constitutively active Cdc42 leads to co-recruitment of the guanine nucleotide exchange factor (GEF) Scd1 and endogenous Cdc42, in a manner dependent on the scaffold protein Scd2. We show that Scd2 function is dispensable when the positive feedback operates through an engineered interaction between the GEF and a Cdc42 effector, the p21-activated kinase 1 (Pak1). Remarkably, this rewired positive feedback confers viability and allows cells to form 2 zones of active Cdc42 even when otherwise essential Cdc42 activators are lacking. These cells further revealed that the small GTPase Ras1 plays a role in both localizing the GEF Scd1 and promoting its activity, which potentiates the positive feedback. We conclude that scaffold-mediated positive feedback, gated by Ras activity, confers robust polarization for rod-shape formation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
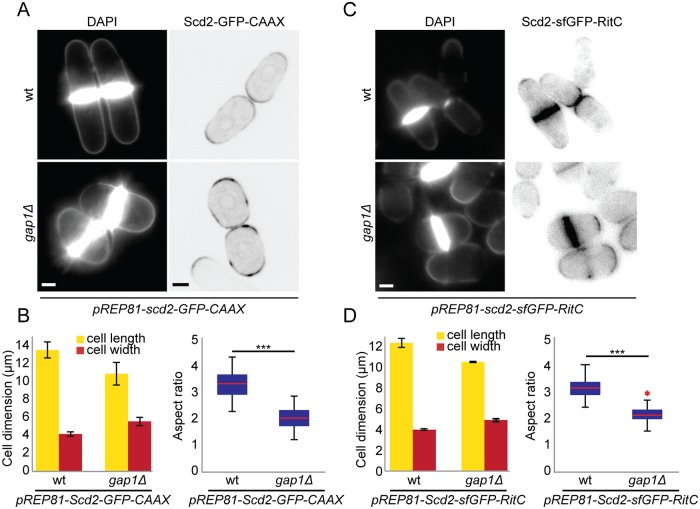
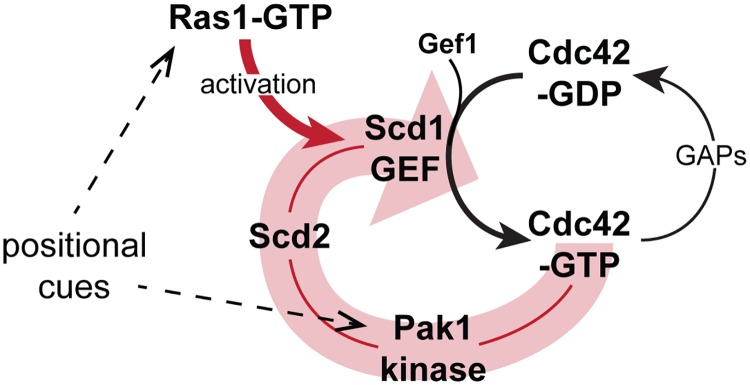
Similar articles
-
Activation of Cdc42 GTPase upon CRY2-Induced Cortical Recruitment Is Antagonized by GAPs in Fission Yeast.Cells. 2020 Sep 12;9(9):2089. doi: 10.3390/cells9092089. Cells. 2020. PMID: 32932721 Free PMC article.
-
A novel interplay between GEFs orchestrates Cdc42 activity during cell polarity and cytokinesis in fission yeast.J Cell Sci. 2019 Dec 3;132(23):jcs236018. doi: 10.1242/jcs.236018. J Cell Sci. 2019. PMID: 31719163 Free PMC article.
-
Eng2, a new player involved in feedback loop regulation of Cdc42 activity in fission yeast.Sci Rep. 2021 Sep 9;11(1):17872. doi: 10.1038/s41598-021-97311-6. Sci Rep. 2021. PMID: 34504165 Free PMC article.
-
Spatial regulation of Cdc42 during cytokinesis.Cell Cycle. 2007 Jul 15;6(14):1687-91. doi: 10.4161/cc.6.14.4481. Epub 2007 May 21. Cell Cycle. 2007. PMID: 17637568 Review.
-
Spontaneous cell polarization: Feedback control of Cdc42 GTPase breaks cellular symmetry.Bioessays. 2015 Nov;37(11):1193-201. doi: 10.1002/bies.201500077. Epub 2015 Sep 4. Bioessays. 2015. PMID: 26338468 Review.
Cited by
-
Redundancy and the role of protein copy numbers in the cell polarization machinery of budding yeast.Nat Commun. 2023 Oct 16;14(1):6504. doi: 10.1038/s41467-023-42100-0. Nat Commun. 2023. PMID: 37845215 Free PMC article.
-
Cdc42 mobility and membrane flows regulate fission yeast cell shape and survival.Nat Commun. 2024 Sep 27;15(1):8363. doi: 10.1038/s41467-024-52655-1. Nat Commun. 2024. PMID: 39333500 Free PMC article.
-
How cells determine the number of polarity sites.Elife. 2021 Apr 26;10:e58768. doi: 10.7554/eLife.58768. Elife. 2021. PMID: 33899733 Free PMC article.
-
Cdc42 reactivation at growth sites is regulated by local cell-cycle-dependent loss of its GTPase-activating protein Rga4 in fission yeast.J Cell Sci. 2021 Oct 1;134(19):jcs259291. doi: 10.1242/jcs.259291. Epub 2021 Oct 11. J Cell Sci. 2021. PMID: 34523683 Free PMC article.
-
Chaperone-Dependent Degradation of Cdc42 Promotes Cell Polarity and Shields the Protein from Aggregation.Mol Cell Biol. 2023;43(5):200-222. doi: 10.1080/10985549.2023.2198171. Epub 2023 Apr 28. Mol Cell Biol. 2023. PMID: 37114947 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
