

The Potyviruses: An Evolutionary Synthesis Is Emerging
- PMID: 31979056
- PMCID: PMC7077269
- DOI: 10.3390/v12020132
The Potyviruses: An Evolutionary Synthesis Is Emerging
Abstract
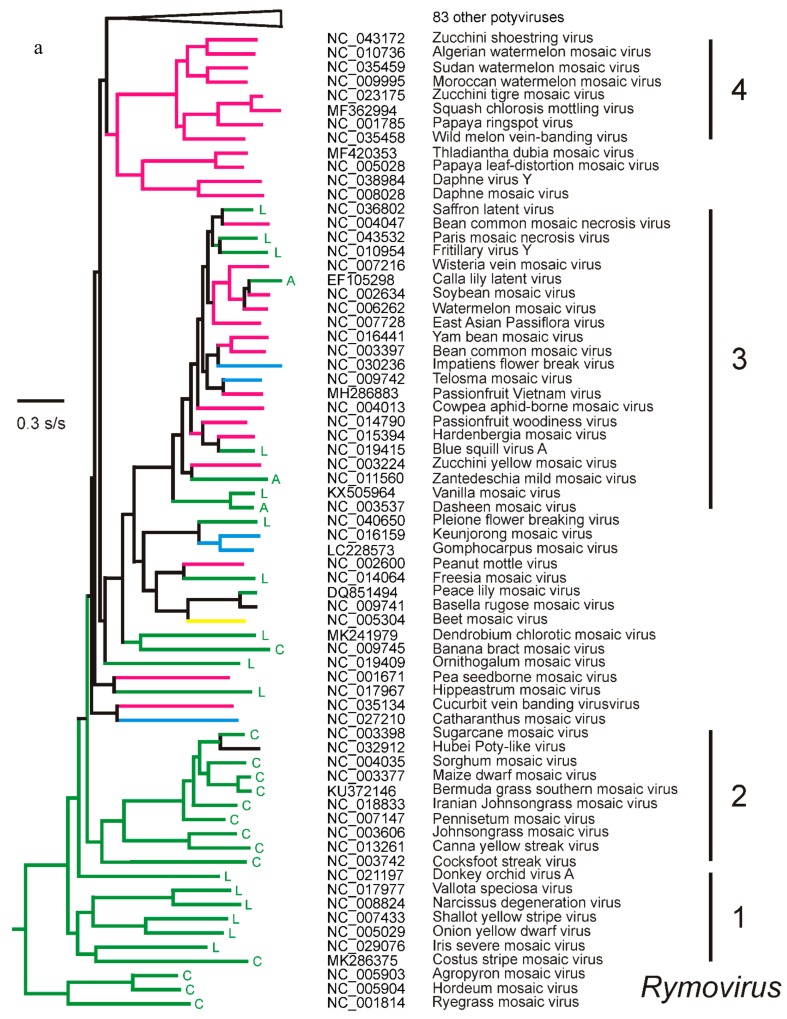
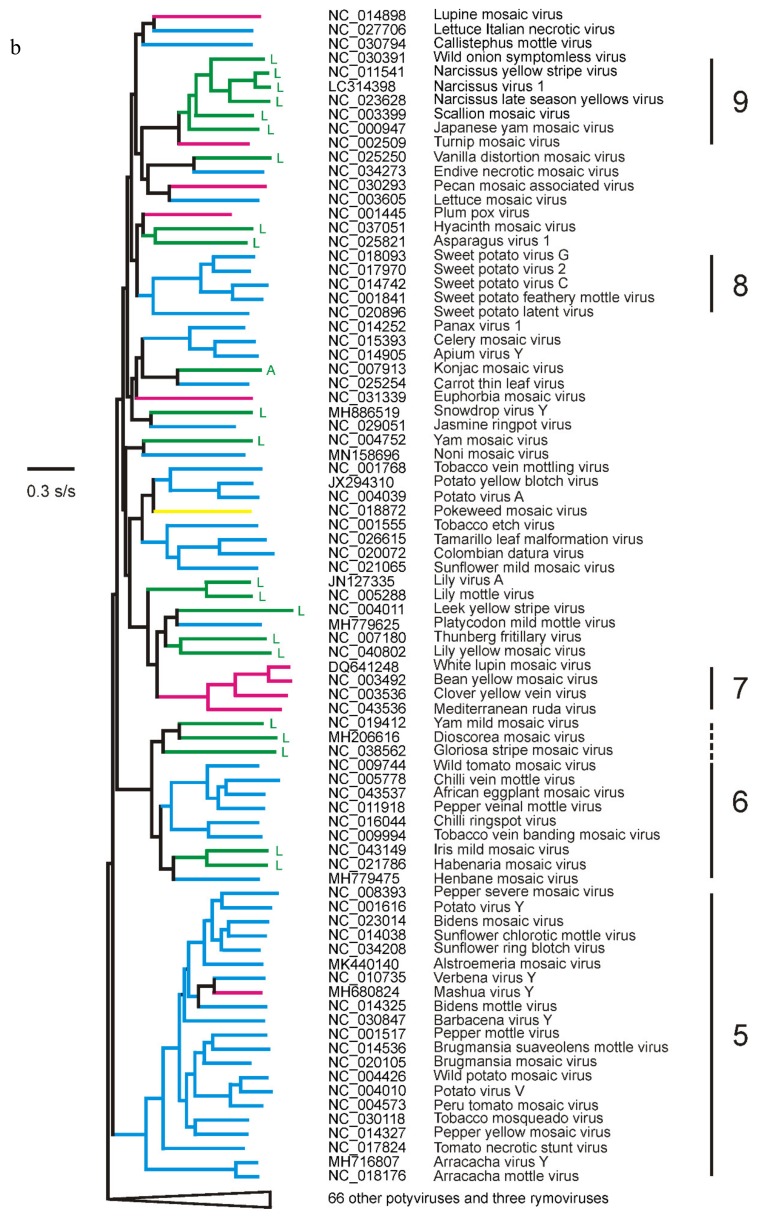
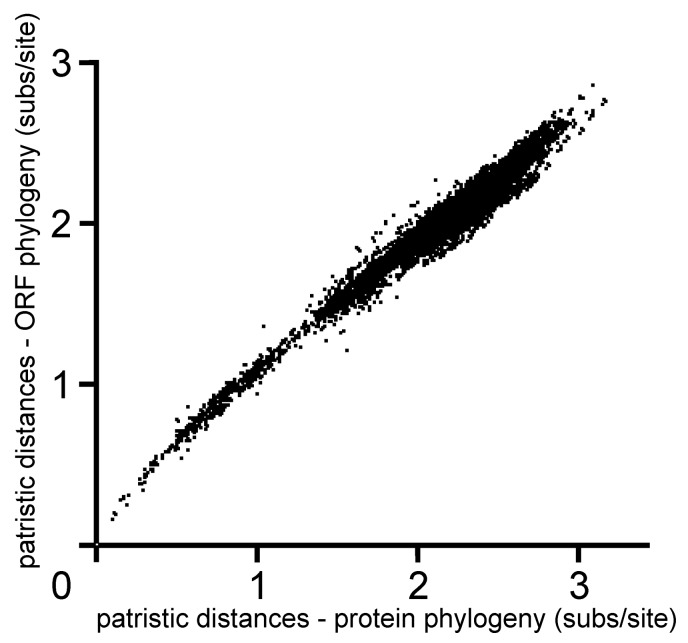
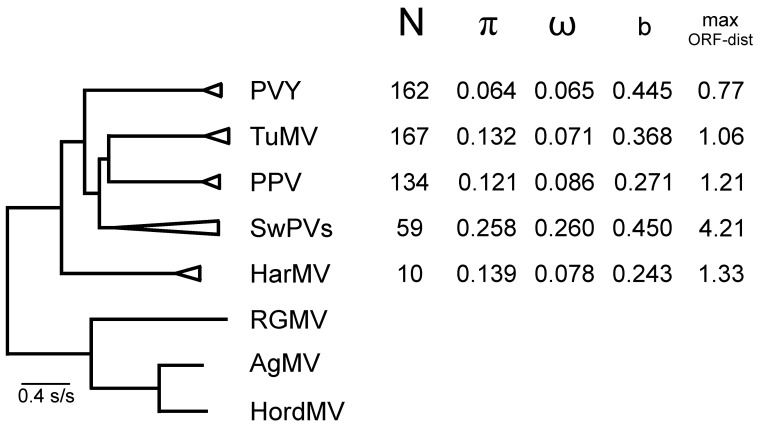
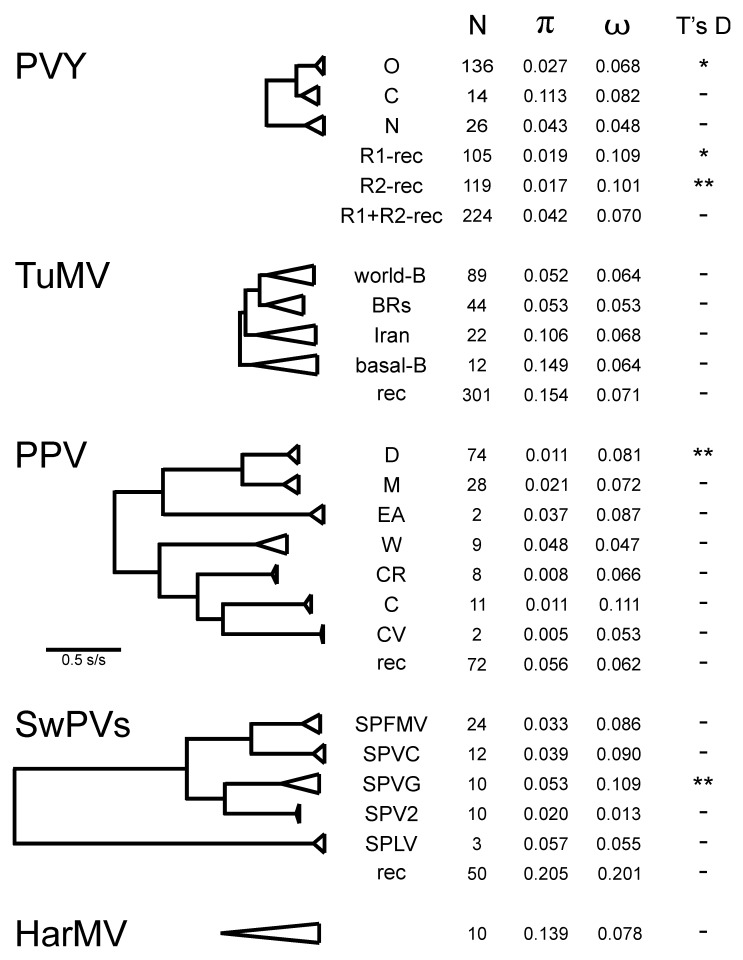
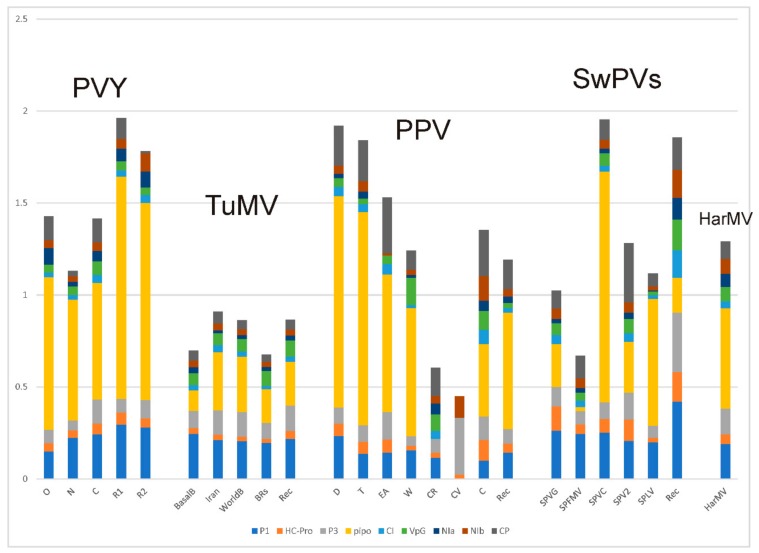
In this review, encouraged by the dictum of Theodosius Dobzhansky that "Nothing in biology makes sense except in the light of evolution", we outline the likely evolutionary pathways that have resulted in the observed similarities and differences of the extant molecules, biology, distribution, etc. of the potyvirids and, especially, its largest genus, the potyviruses. The potyvirids are a family of plant-infecting RNA-genome viruses. They had a single polyphyletic origin, and all share at least three of their genes (i.e., the helicase region of their CI protein, the RdRp region of their NIb protein and their coat protein) with other viruses which are otherwise unrelated. Potyvirids fall into 11 genera of which the potyviruses, the largest, include more than 150 distinct viruses found worldwide. The first potyvirus probably originated 15,000-30,000 years ago, in a Eurasian grass host, by acquiring crucial changes to its coat protein and HC-Pro protein, which enabled it to be transmitted by migrating host-seeking aphids. All potyviruses are aphid-borne and, in nature, infect discreet sets of monocotyledonous or eudicotyledonous angiosperms. All potyvirus genomes are under negative selection; the HC-Pro, CP, Nia, and NIb genes are most strongly selected, and the PIPO gene least, but there are overriding virus specific differences; for example, all turnip mosaic virus genes are more strongly conserved than those of potato virus Y. Estimates of dN/dS (ω) indicate whether potyvirus populations have been evolving as one or more subpopulations and could be used to help define species boundaries. Recombinants are common in many potyvirus populations (20%-64% in five examined), but recombination seems to be an uncommon speciation mechanism as, of 149 distinct potyviruses, only two were clear recombinants. Human activities, especially trade and farming, have fostered and spread both potyviruses and their aphid vectors throughout the world, especially over the past five centuries. The world distribution of potyviruses, especially those found on islands, indicates that potyviruses may be more frequently or effectively transmitted by seed than experimental tests suggest. Only two meta-genomic potyviruses have been recorded from animal samples, and both are probably contaminants.
Keywords: evolution; metagenomes; phylogenetics; population genetics; potyvirids; potyviruses; recombination; speciation.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures


Similar articles
-
Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus.Viruses. 2020 Jul 17;12(7):773. doi: 10.3390/v12070773. Viruses. 2020. PMID: 32708998 Free PMC article. Review.
-
Turnip mosaic potyvirus probably first spread to Eurasian brassica crops from wild orchids about 1000 years ago.PLoS One. 2013;8(2):e55336. doi: 10.1371/journal.pone.0055336. Epub 2013 Feb 6. PLoS One. 2013. PMID: 23405136 Free PMC article.
-
The potyviruses of Australia.Arch Virol. 2008;153(8):1411-20. doi: 10.1007/s00705-008-0134-6. Epub 2008 Jun 20. Arch Virol. 2008. PMID: 18566735
-
Engineering aphid transmission of foxtail mosaic virus in the presence of potyvirus helper component proteinase through coat protein modifications.J Gen Virol. 2023 Apr;104(4). doi: 10.1099/jgv.0.001844. J Gen Virol. 2023. PMID: 37053090
-
Molecular biology of potyviruses.Adv Virus Res. 2015;92:101-99. doi: 10.1016/bs.aivir.2014.11.006. Epub 2015 Jan 21. Adv Virus Res. 2015. PMID: 25701887 Review.
Cited by
-
The P1 Protein of Watermelon mosaic virus Compromises the Activity as RNA Silencing Suppressor of the P25 Protein of Cucurbit yellow stunting disorder virus.Front Microbiol. 2021 Mar 22;12:645530. doi: 10.3389/fmicb.2021.645530. eCollection 2021. Front Microbiol. 2021. PMID: 33828542 Free PMC article.
-
Pepper Mottle Virus and Its Host Interactions: Current State of Knowledge.Viruses. 2021 Sep 25;13(10):1930. doi: 10.3390/v13101930. Viruses. 2021. PMID: 34696360 Free PMC article. Review.
-
Ecological Strategies for Resource Use by Three Bromoviruses in Anthropic and Wild Plant Communities.Viruses. 2023 Aug 21;15(8):1779. doi: 10.3390/v15081779. Viruses. 2023. PMID: 37632121 Free PMC article.
-
Multiple Infections with Viruses of the Family Tymoviridae in Czech Grapevines.Viruses. 2024 Feb 23;16(3):343. doi: 10.3390/v16030343. Viruses. 2024. PMID: 38543709 Free PMC article.
-
Enhanced Apiaceous Potyvirus Phylogeny, Novel Viruses, and New Country and Host Records from Sequencing Apiaceae Samples.Plants (Basel). 2022 Jul 27;11(15):1951. doi: 10.3390/plants11151951. Plants (Basel). 2022. PMID: 35956429 Free PMC article.
References
-
- Salaman R.N. Degeneration of the potato. An urgent problem. J. Nat. Inst. Agric. Bot. 1925;1:30–51.
-
- Smith K.M. On the composite nature of certain potato virus diseases of the mosaic group as revealed by the use of plant indicators and selective methods of transmission. Proc. Roy. Soc. Ser. B. 1931;109:251–266. doi: 10.1098/rspb.1931.0080. - DOI
-
- Hamilton M.A. On three new viruses diseases in Hyoscyamus niger. Ann. Appl. Biol. 1932;19:550–567. doi: 10.1111/j.1744-7348.1932.tb04341.x. - DOI
-
- Barnett O.W. A summary of potyvirus taxonomy and definitions. In: Barnett O.W., editor. Potyvirus Taxonomy. Volume 5. Springer; Vienna, Austria: 1992. pp. 435–444. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
