

Validation of a New Multicistronic Plasmid for the Efficient and Stable Expression of Transgenes in Microalgae
- PMID: 31979077
- PMCID: PMC7037629
- DOI: 10.3390/ijms21030718
Validation of a New Multicistronic Plasmid for the Efficient and Stable Expression of Transgenes in Microalgae
Abstract
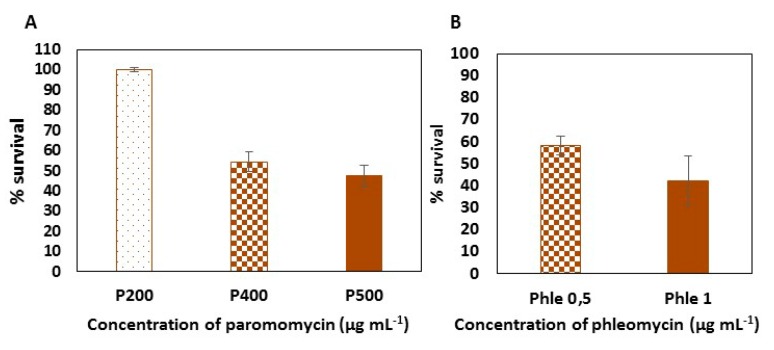
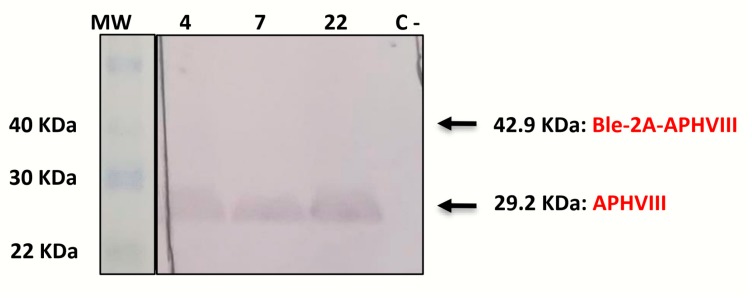
Low stability of transgenes and high variability of their expression levels among the obtained transformants are still pending challenges in the nuclear genetic transformation of microalgae. We have generated a new multicistronic microalgal expression plasmid, called Phyco69, to make easier the large phenotypic screening usually necessary for the selection of high-expression stable clones. This plasmid contains a polylinker region (PLK) where any gene of interest (GOI) can be inserted and get linked, through a short viral self-cleaving peptide to the amino terminus of the aminoglycoside 3'-phosphotransferase (APHVIII) from Streptomyces rimosus, which confers resistance to the antibiotic paromomycin. The plasmid has been validated by expressing a second antibiotic resistance marker, the ShBLE gene, which confers resistance to phleomycin. It has been shown, by RT-PCR and by phenotypic studies, that the fusion of the GOI to the selective marker gene APHVIII provides a simple method to screen and select the transformants with the highest level of expression of both the APHVIII gene and the GOI among the obtained transformants. Immunodetection studies have shown that the multicistronic transcript generated from Phyco69 is correctly processed, producing independent gene products from a common promoter.
Keywords: 2A; microalgae transformation; multicistronic transcript.; paromomycin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
