

Regulation of adipose tissue inflammation by interleukin 6
- PMID: 31980524
- PMCID: PMC7022151
- DOI: 10.1073/pnas.1920004117
Regulation of adipose tissue inflammation by interleukin 6
Abstract
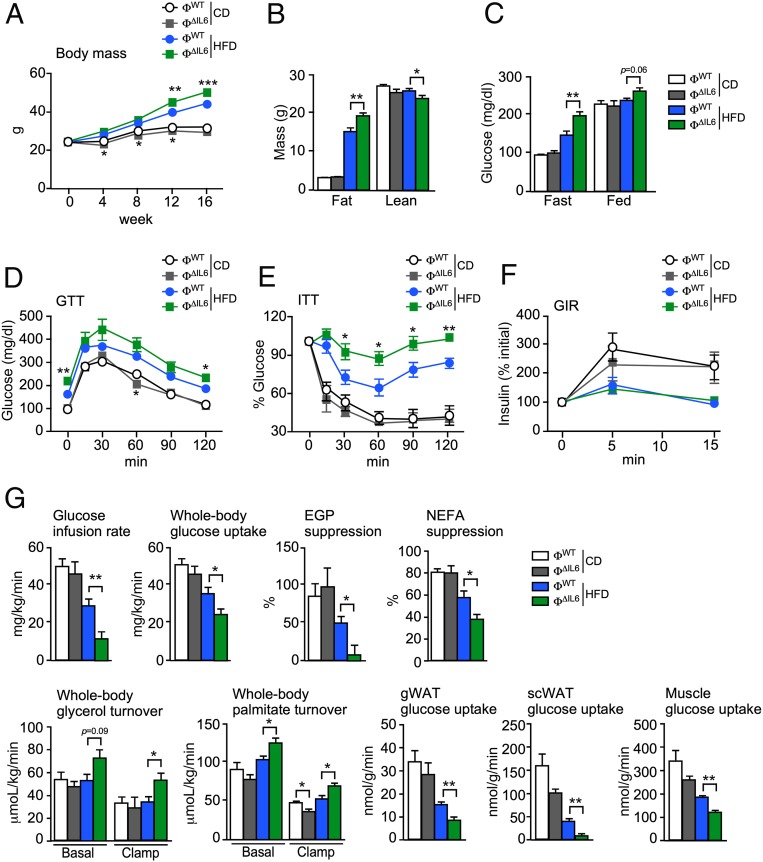
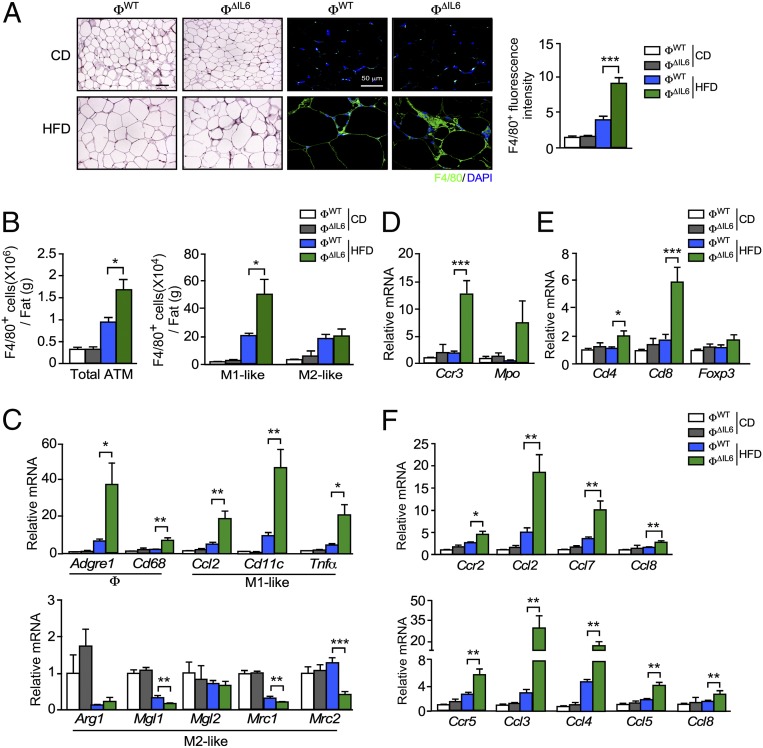
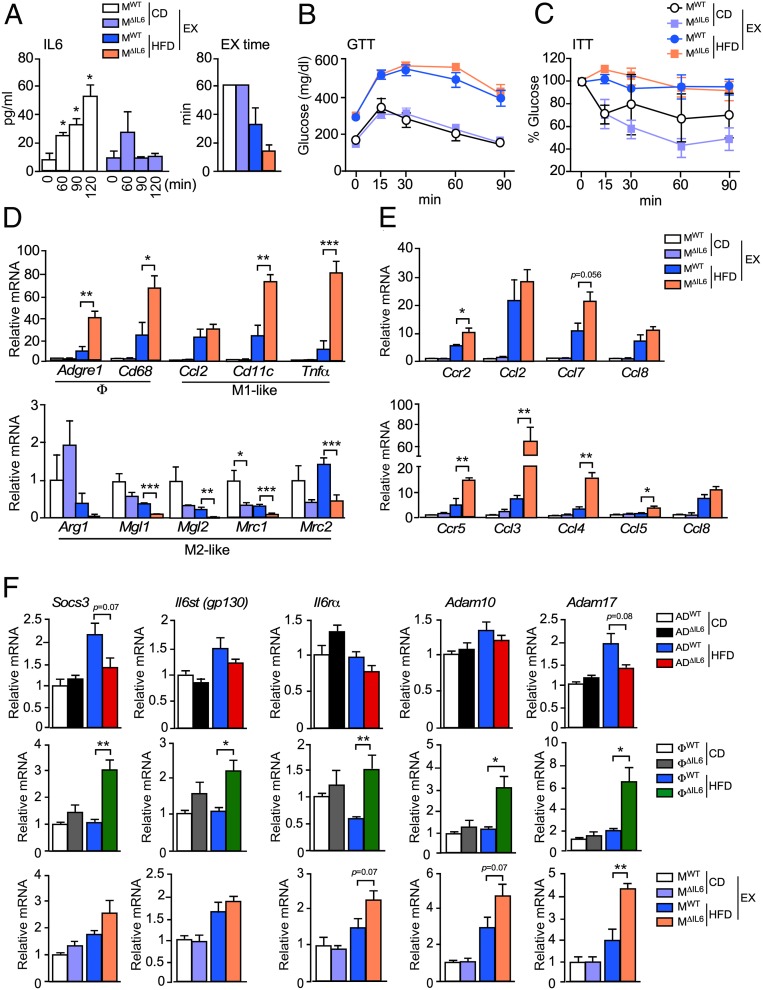
Obesity is associated with a chronic state of low-grade inflammation and progressive tissue infiltration by immune cells and increased expression of inflammatory cytokines. It is established that interleukin 6 (IL6) regulates multiple aspects of metabolism, including glucose disposal, lipolysis, oxidative metabolism, and energy expenditure. IL6 is secreted by many tissues, but the role of individual cell types is unclear. We tested the role of specific cells using a mouse model with conditional expression of the Il6 gene. We found that IL6 derived from adipocytes increased, while IL6 derived from myeloid cells and muscle suppressed, macrophage infiltration of adipose tissue. These opposite actions were associated with a switch of IL6 signaling from a canonical mode (myeloid cells) to a noncanonical trans-signaling mode (adipocytes and muscle) with increased expression of the ADAM10/17 metalloprotease that promotes trans-signaling by the soluble IL6 receptor α. Collectively, these data demonstrate that the source of IL6 production plays a major role in the physiological regulation of metabolism.
Keywords: adipose tissue; inflammation; insulin resistance; interleukin 6.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
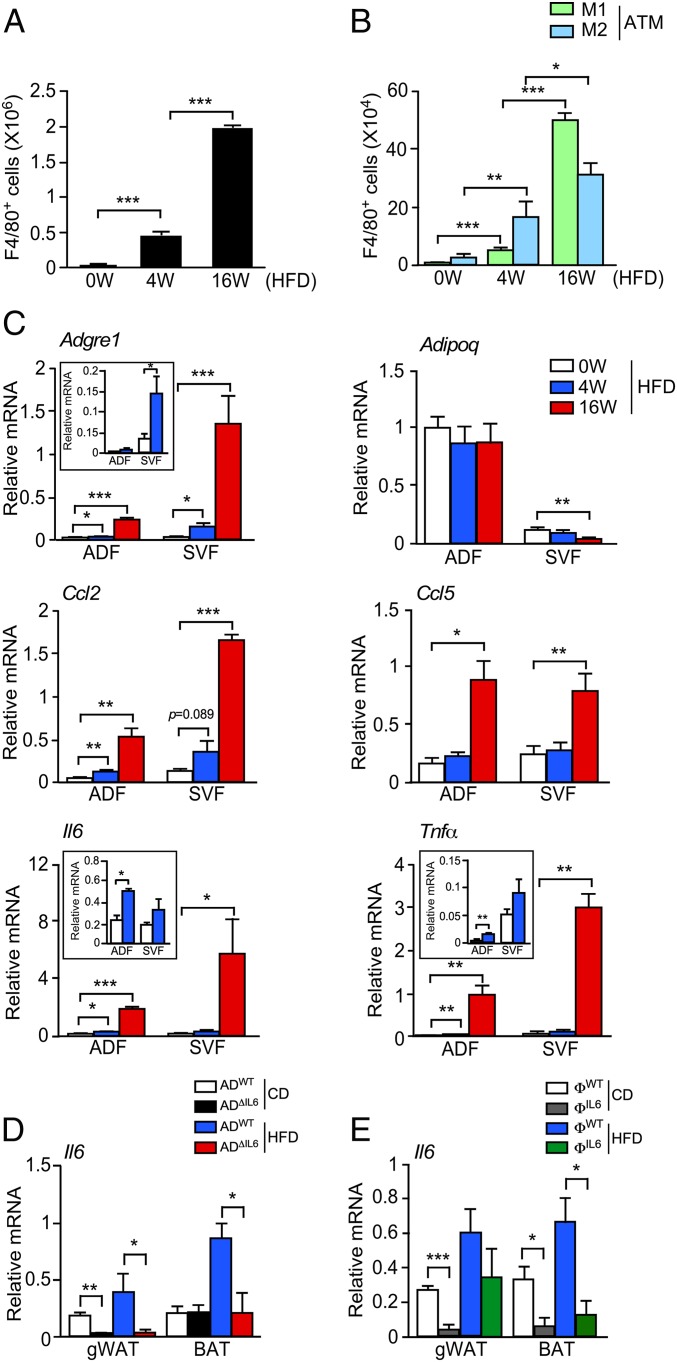
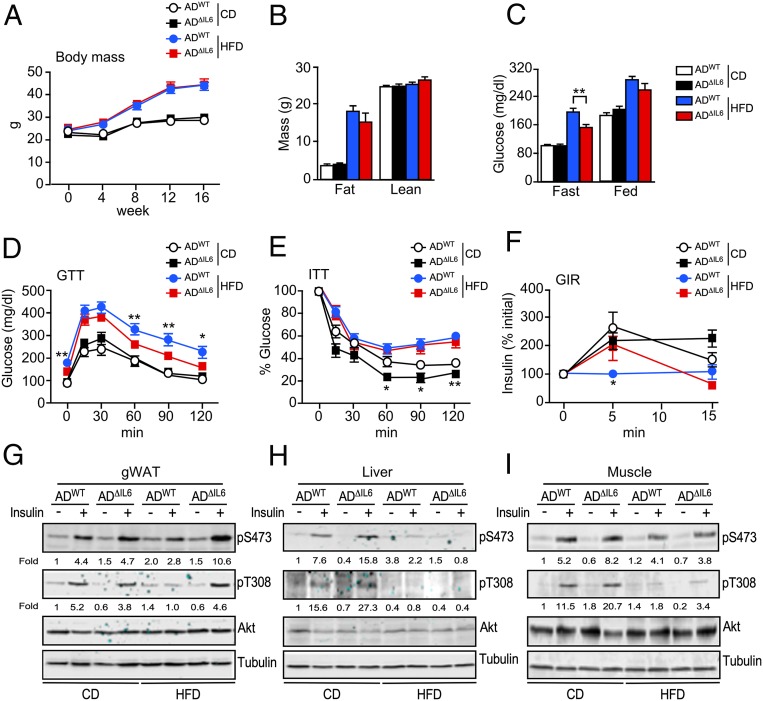
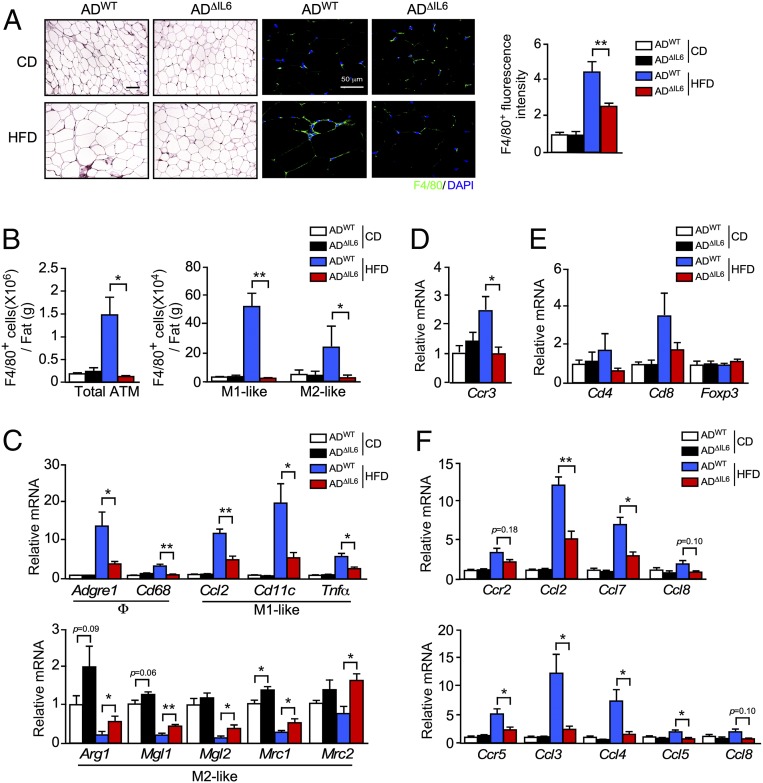
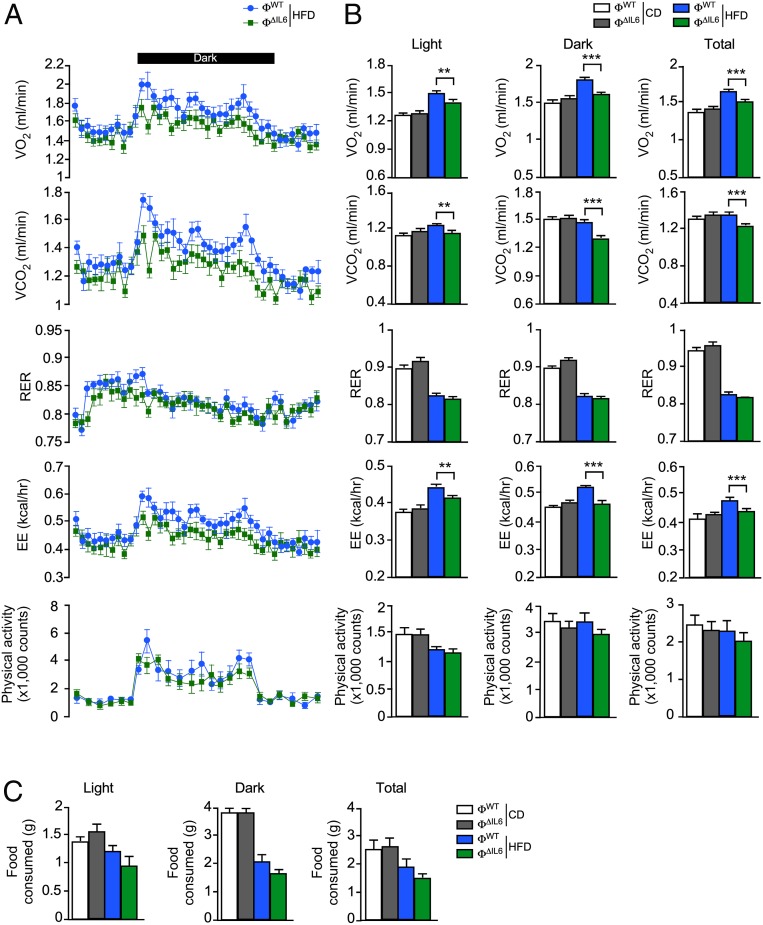
References
-
- Mauer J., Denson J. L., Brüning J. C., Versatile functions for IL-6 in metabolism and cancer. Trends Immunol. 36, 92–101 (2015). - PubMed
-
- Carey A. L., et al. , Interleukin-6 increases insulin-stimulated glucose disposal in humans and glucose uptake and fatty acid oxidation in vitro via AMP-activated protein kinase. Diabetes 55, 2688–2697 (2006). - PubMed
-
- van Hall G., et al. , Interleukin-6 stimulates lipolysis and fat oxidation in humans. J. Clin. Endocrinol. Metab. 88, 3005–3010 (2003). - PubMed
-
- Schmidt-Arras D., Rose-John S., IL-6 pathway in the liver: From physiopathology to therapy. J. Hepatol. 64, 1403–1415 (2016). - PubMed
-
- Scheller J., Chalaris A., Schmidt-Arras D., Rose-John S., The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 1813, 878–888 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
