

Tissue specificity of senescent cell accumulation during physiologic and accelerated aging of mice
- PMID: 31981461
- PMCID: PMC7059165
- DOI: 10.1111/acel.13094
Tissue specificity of senescent cell accumulation during physiologic and accelerated aging of mice
Abstract
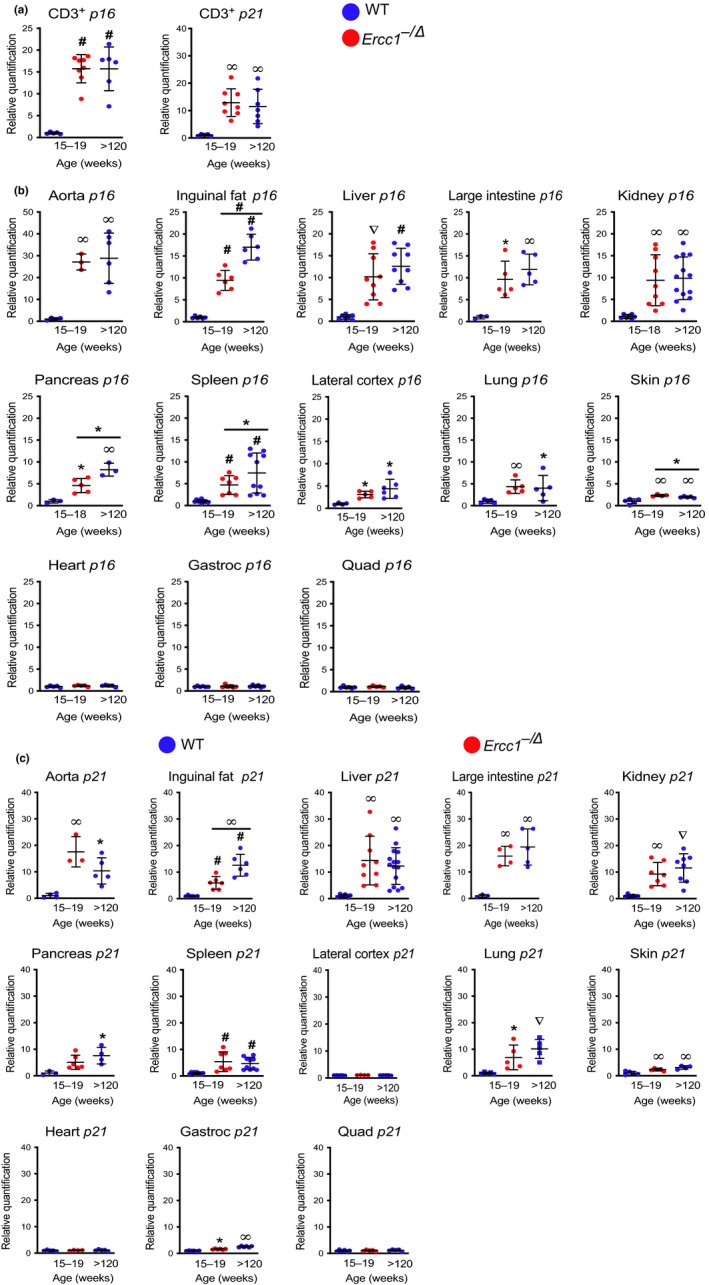
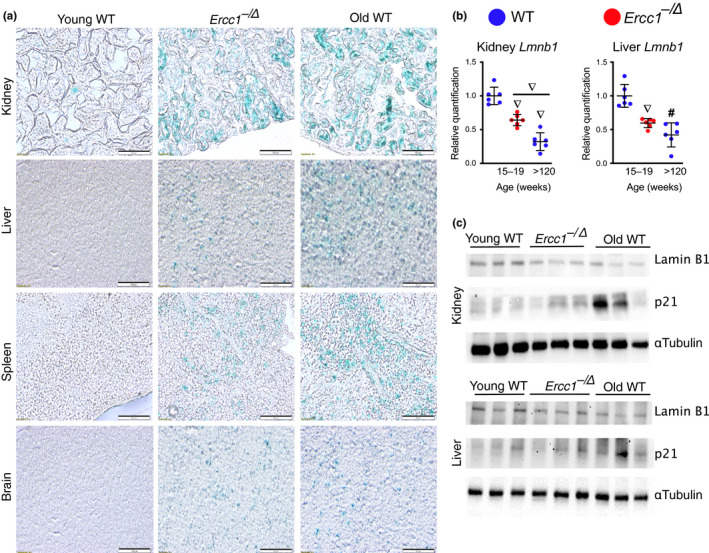
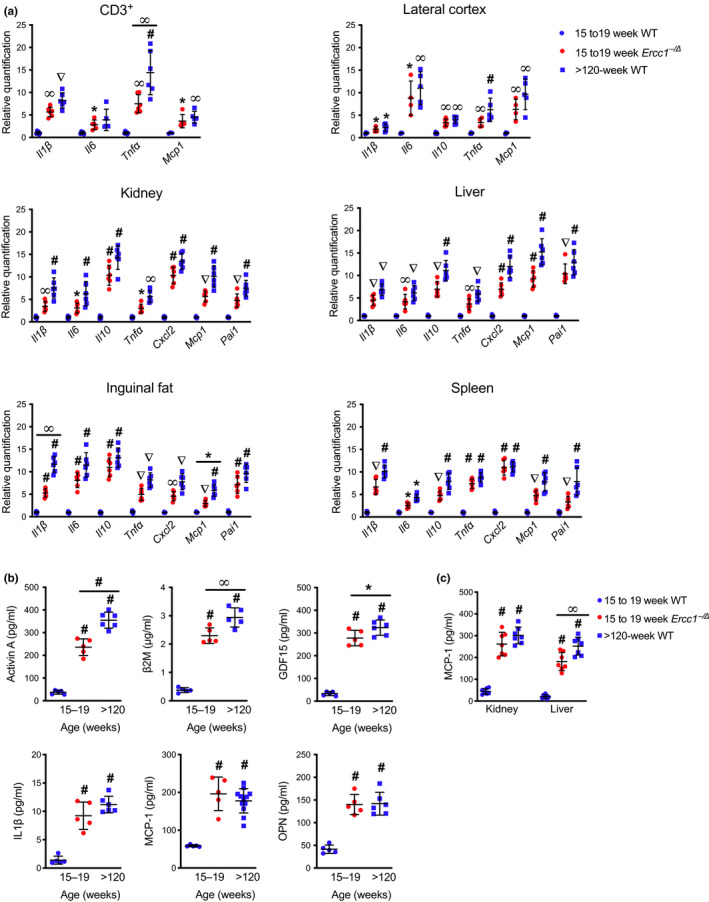
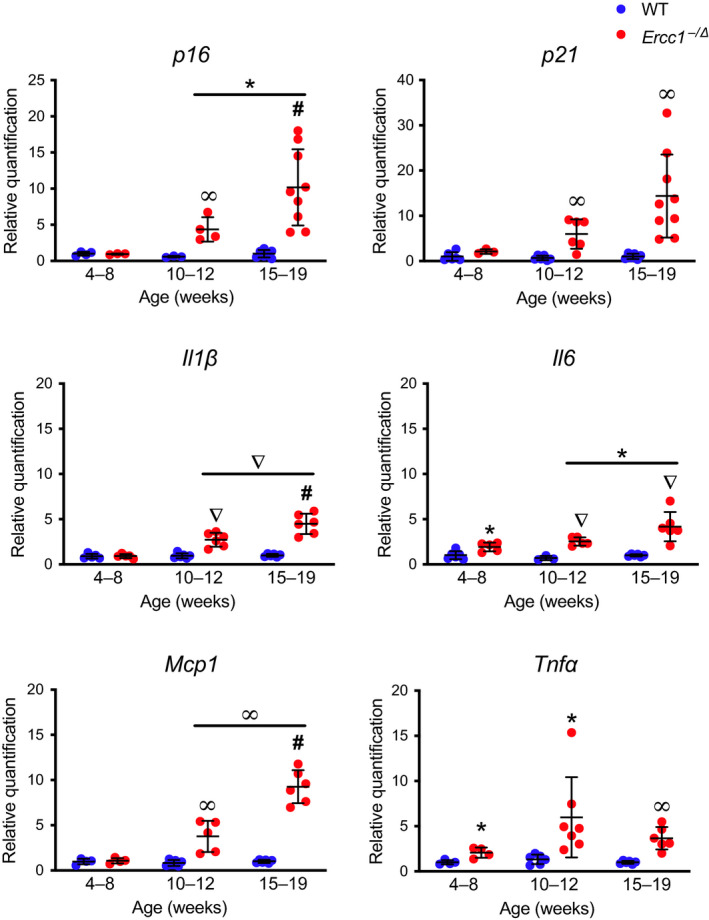
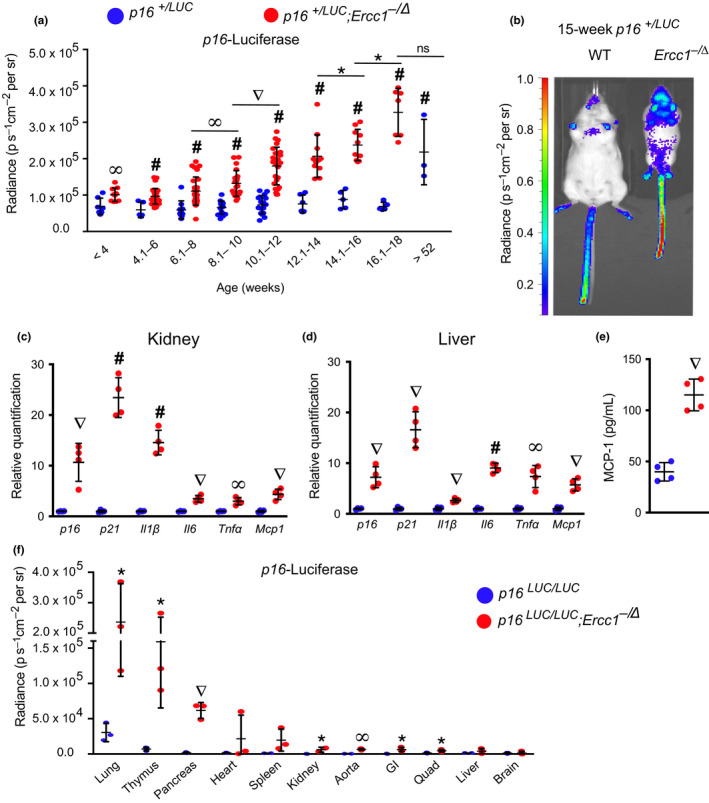
Senescent cells accumulate with age in vertebrates and promote aging largely through their senescence-associated secretory phenotype (SASP). Many types of stress induce senescence, including genotoxic stress. ERCC1-XPF is a DNA repair endonuclease required for multiple DNA repair mechanisms that protect the nuclear genome. Humans or mice with reduced expression of this enzyme age rapidly due to increased levels of spontaneous, genotoxic stress. Here, we asked whether this corresponds to an increased level of senescent cells. p16Ink4a and p21Cip1 mRNA were increased ~15-fold in peripheral lymphocytes from 4- to 5-month-old Ercc1-/∆ and 2.5-year-old wild-type (WT) mice, suggesting that these animals exhibit a similar biological age. p16Ink4a and p21Cip1 mRNA were elevated in 10 of 13 tissues analyzed from 4- to 5-month-old Ercc1-/∆ mice, indicating where endogenous DNA damage drives senescence in vivo. Aged WT mice had similar increases of p16Ink4a and p21Cip1 mRNA in the same 10 tissues as the mutant mice. Senescence-associated β-galactosidase activity and p21Cip1 protein also were increased in tissues of the progeroid and aged mice, while Lamin B1 mRNA and protein levels were diminished. In Ercc1-/Δ mice with a p16Ink4a luciferase reporter, bioluminescence rose steadily with age, particularly in lung, thymus, and pancreas. These data illustrate where senescence occurs with natural and accelerated aging in mice and the relative extent of senescence among tissues. Interestingly, senescence was greater in male mice until the end of life. The similarities between Ercc1-/∆ and aged WT mice support the conclusion that the DNA repair-deficient mice accurately model the age-related accumulation of senescent cells, albeit six-times faster.
Keywords: DNA repair; ERCC1-XPF; aging; cellular senescence; endogenous DNA damage; progeria.
© 2020 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
None declared.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
