

Copper modulates sex-specific fructose hepatoxicity in nonalcoholic fatty liver disease (NALFD) Wistar rat models
- PMID: 31986483
- PMCID: PMC7078045
- DOI: 10.1016/j.jnutbio.2019.108316
Copper modulates sex-specific fructose hepatoxicity in nonalcoholic fatty liver disease (NALFD) Wistar rat models
Abstract
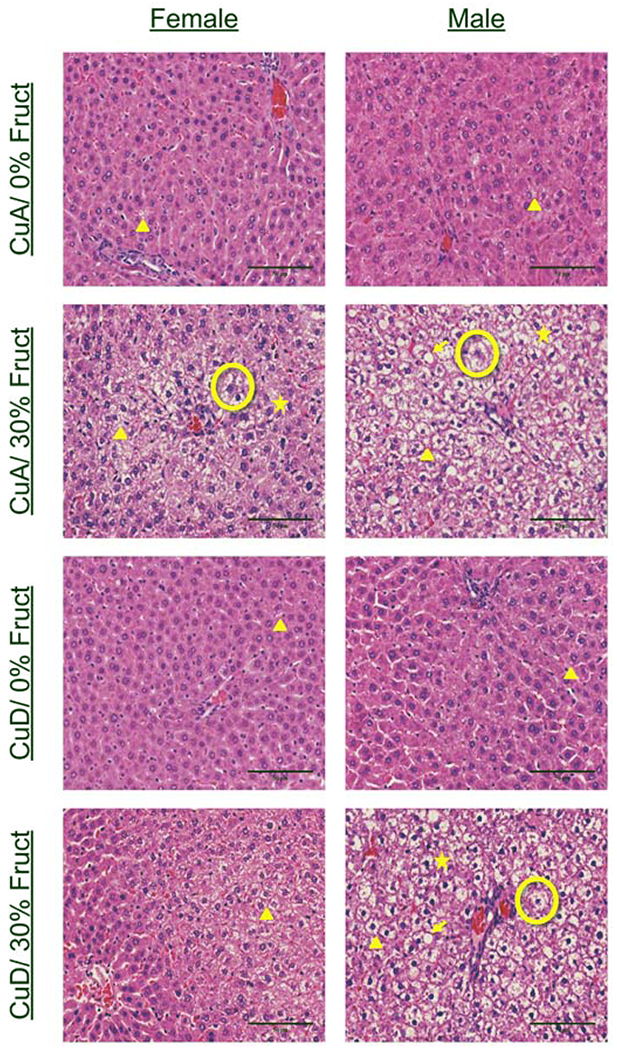
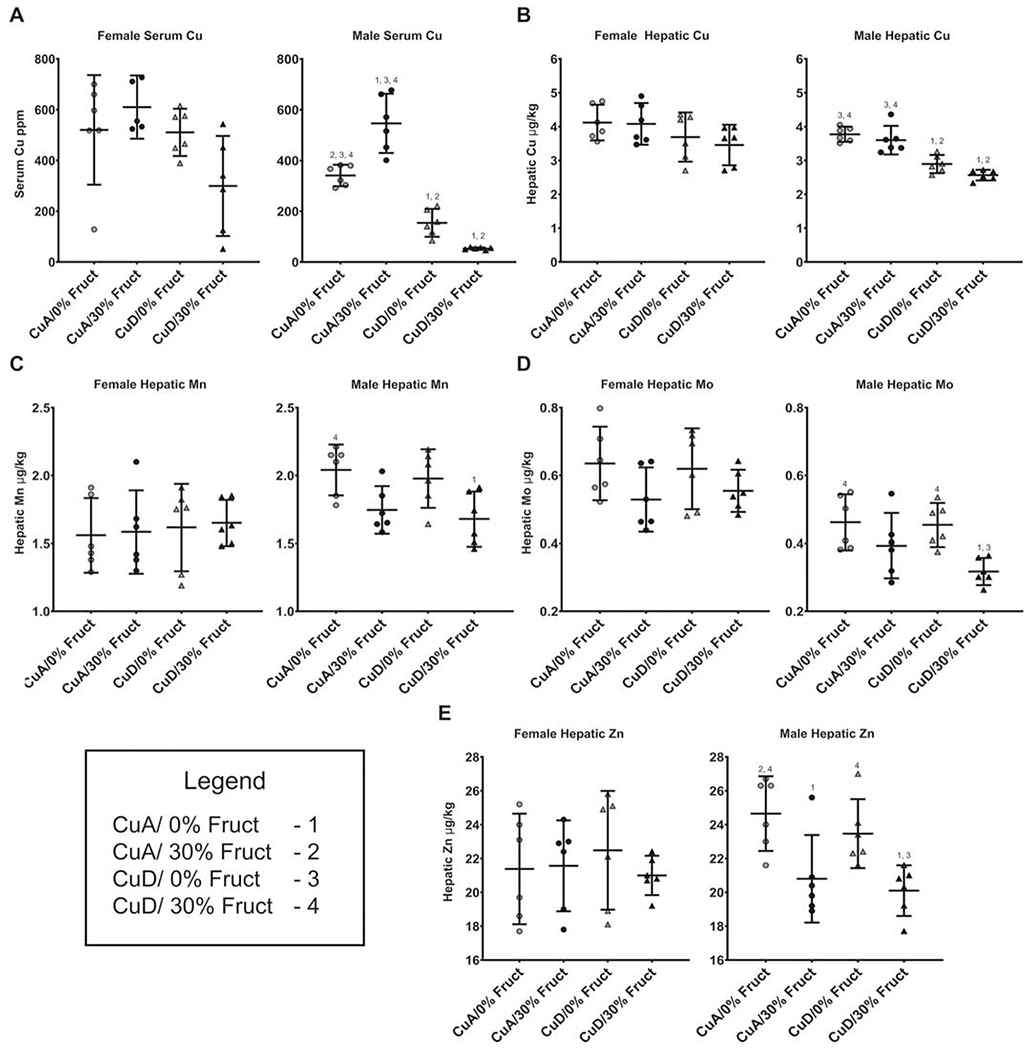
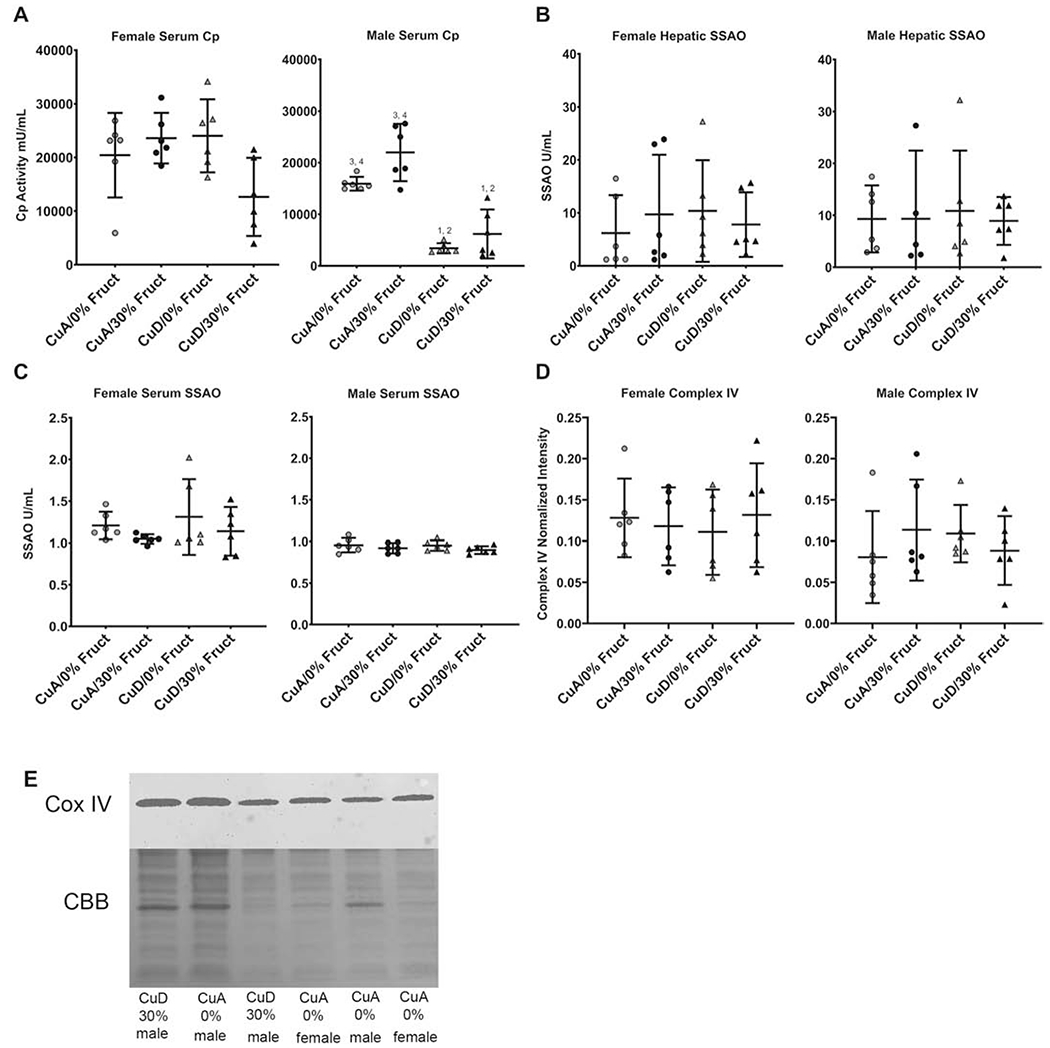
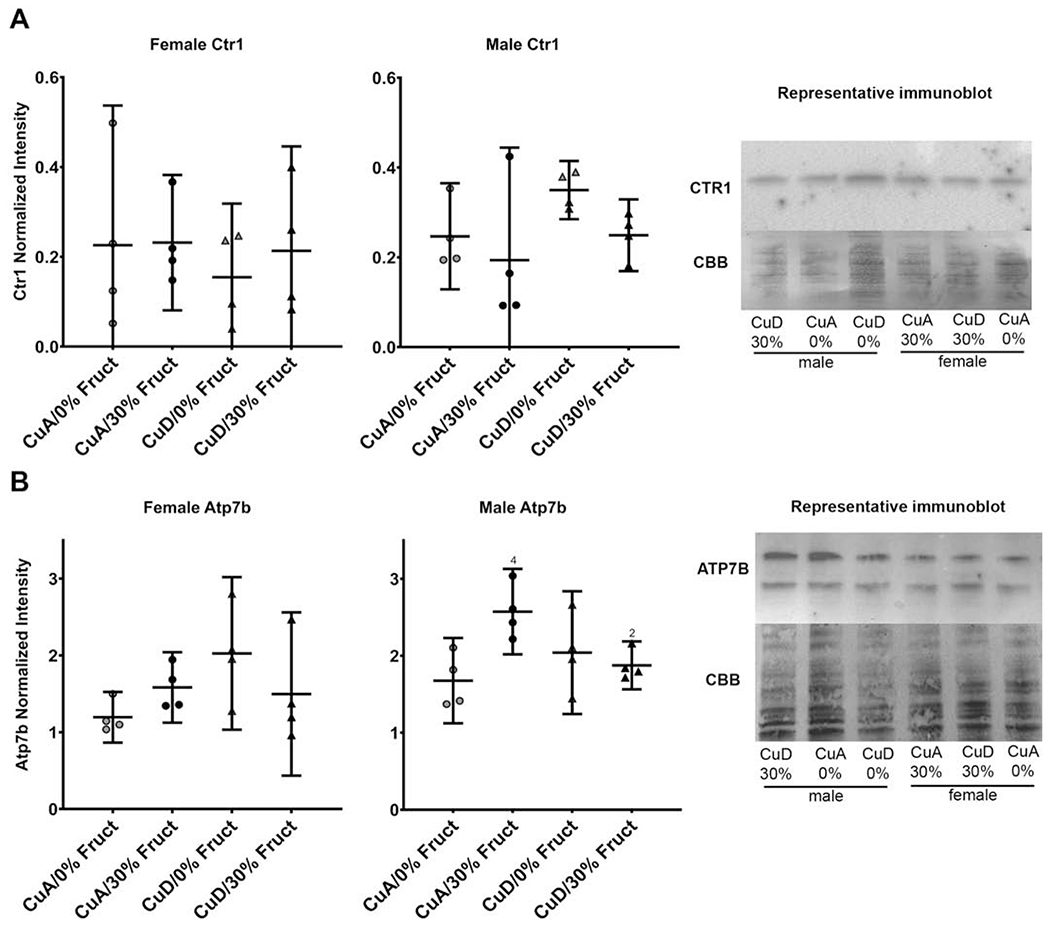
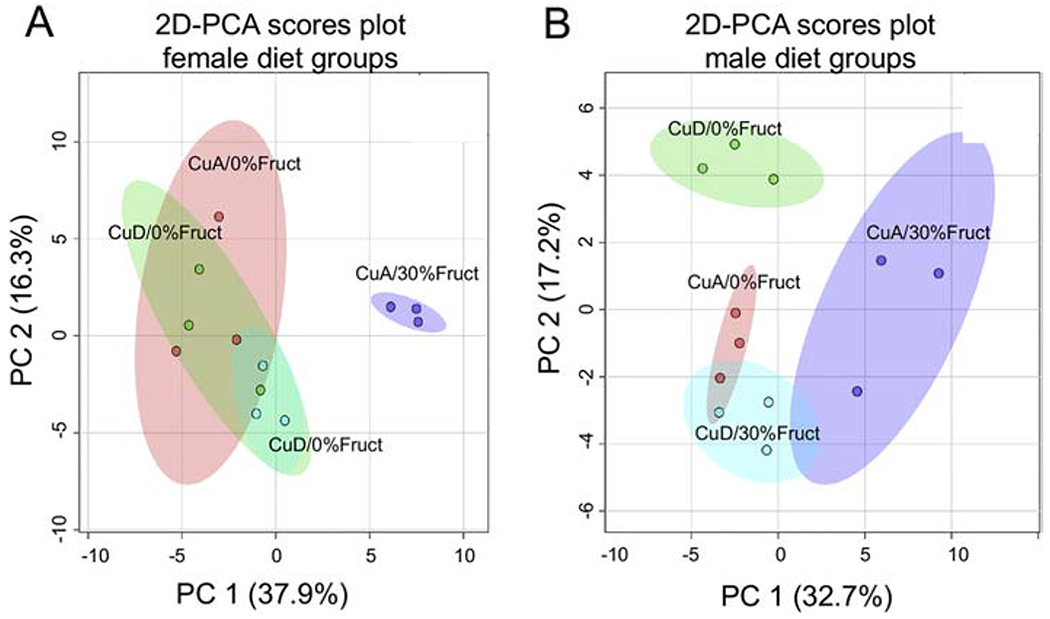
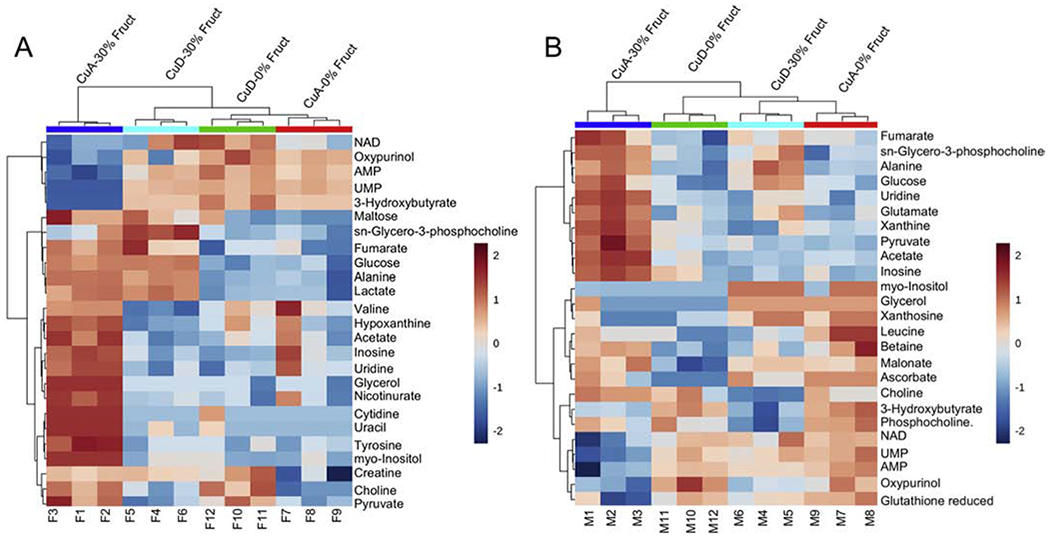
This study aimed to characterize the impact of dietary copper on the biochemical and hepatic metabolite changes associated with fructose toxicity in a Wistar rat model of fructose-induced liver disease. Twenty-four male and 24 female, 6-week-old, Wister rats were separated into four experimental dietary treatment groups (6 males and 6 females per group), as follows: (1) a control diet: containing no fructose with adequate copper (i.e., CuA/0% Fruct); (2) a diet regimen identical to the control and supplemented with 30% w/v fructose in the animals' drinking water (CuA/30% Fruct); (3) a diet identical to the control diet but deficient in copper content (CuD/0% Fruct) and (4) a diet identical to the control diet but deficient in copper content and supplemented with 30% w/v fructose in the drinking water (CuD/30% Fruct). The animals were fed the four diet regimens for 5 weeks, followed by euthanization and assessment of histology, elemental profiles and identification and quantitation of liver metabolites. Results from 1H nuclear magnetic resonance metabolomics revealed mechanistic insights into copper modulation of fructose hepatotoxicity through identification of distinct metabolic phenotypes that were highly correlated with diet and sex. This study also identified previously unknown sex-specific responses to both fructose supplementation and restricted copper intake, while the presence of adequate dietary copper promoted most pronounced fructose-induced metabolite changes.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures
References
-
- NHANES - National Health and Nutrition Examination Survey Homepage n.d. https://www.cdc.gov/nchs/nhanes/ (accessed January 24, 2017).
-
- Tang Z, Gasperkova D, Xu J, Baillie R, Lee JH, Clarke SD. Copper deficiency induces hepatic fatty acid synthase gene transcription in rats by increasing the nuclear content of mature sterol regulatory element binding protein 1. J Nutr 2000;130:2915–21. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
